Vision
The eye is an elaborate organ whose primary function is to collect and focus light upon the photosensitive retina. It lies within a coneshaped cavity of the skull, the orbit, which houses the eyeball (globe) and a number of other soft tissue structures, the ocular adnexa (e.g., muscles, glands), that act upon the eyeball in service of its light-collecting function.
Unlike the human orbit, which is a complete bony cone, the ventral portion of the orbit of domestic species is bounded by soft tissues, notably the pterygoid muscles.Ocular Adnexa
Palpebrae. Two mobile folds of haired skin protect the anterior aspect of the eyeball. These are the eyelids, or palpebrae. The gap between the margins of the two palpebrae is the palpebral fissure, and its shape and size are controlled by the muscles of the eyelids. A layer of dense connective tissue in the palpebrae, the tarsal plate (tarsus), affords them a certain amount of rigidity. The palpebrae have thin but otherwise typical haired skin on their superficial side. This skin ends at the lid margin with a sharp transition to mucous membrane. The margin may have cilia (eyelashes).
Abundant modified sweat and sebaceous glands are associated with the lid margin. A row of large modified sebaceous glands, the tarsal glands (meibomian glands), is especially clinically significant, as they commonly become infected, develop tumors, or become impacted. The tarsal glands are present in both palpebrae and open into a shallow furrow near the mucocutaneous junction of the eyelid. The openings of their ducts are readily seen when the margin of the lid is everted, and the glands themselves are usually visible as yellowish-white columns under the mucous membrane on the inner surface of the lid. The tarsal glands produce an important oil layer of the tear film.
The conjunctiva is the mucous membrane that lines the inside of the palpebrae and part of the anterior surface of the eyeball (excluding the clear cornea).
Diseases of the conjunctiva are the most common eye disorders. The conjunctiva is rich in mucus-producing glands, lymphocytes, nerves, and blood vessels. The very small space between the eyelids and the surface of the eye is the conjunctival sac, and it is into this space that eye medications are usually instilled.Inflammation of the conjunctiva, or conjunctivitis, is characterized by redness and increased mucus production. Cancer eye, squamous cell carcinoma of the conjunctiva, is relatively common in Hereford, Holstein, and shorthorn breeds of cattle.
Domestic species possess a nictitating membrane, or third eyelid. This is a fold of mucous membrane arising from the ventromedial aspect of the conjunctival sac between the eyeball and palpebrae. It is given rigidity by a T-shaped cartilage within it, and it smooths the tear film and protects the cornea.
The third eyelid has at its base a serous gland, called simply the gland of the third eyelid, which normally contributes about 50% of the tear film. Ruminants, the pig, and laboratory rodents possess another deeper gland associated with the third eyelid. This is the harderian gland. It, too, contributes to the tear film.
Lacrimal Apparatus. The lacrimal apparatus comprises a series of serous, seromucoid, and mucous glands and the duct systems that drain
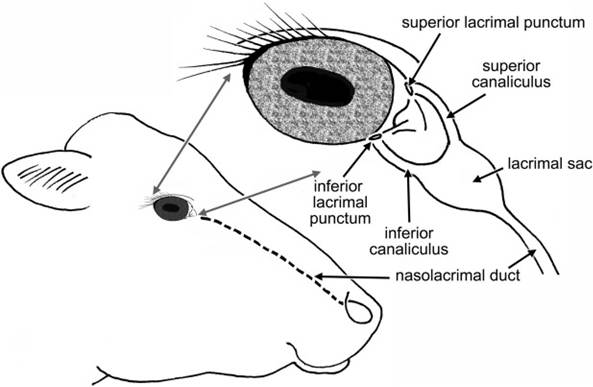
Figure 11-9. The lacrimal apparatus. Tears produced by the lacrimal gland and gland of the third eyelid are drained at the medial corner of the eye at superior and inferior lacrimal puncta. The two canaliculi converge at the lacrimal sac, and from there tears flow via the nasolacrimal duct to the vestibule of the nostril.
their secretions from the conjunctival sac (Fig. 11-9). It provides a moist environment for the anterior surface of the eye. The lacrimal gland lies in the dorsolateral portion of the orbit; its secretion, together with that of the gland of the third eyelid, is the major contributor to the tear film.
In addition to the lacrimal gland and the gland of the third eyelid, the smaller, more diffuse glands of the conjunctiva and the eyelids contribute oil and mucus to the tear film.The tears drain from the conjunctival sac via two small openings on upper and lower palpebrae near the medial corner. These are the superior and inferior lacrimal puncta, which open into short ducts (superior and inferior canaliculi) that join one another at a small sac (lacrimal sac) near the medial corner of the eye that is the origin of the nasolacrimal duct. The nasolacrimal duct travels through the bones of the face to open into the nasal cavity andIor just inside the nostril. Moisture on the nose of domestic animals is largely derived from the tears.
Extraocular Muscles. The globe of the eye moves by the action of seven striated muscles, designated extraocular muscles to distinguish them from the intraocular muscles that lie entirely within the eyeball (Fig. 11-10).
The m. retractor bulbi arises in the caudal part of the orbit, lateral to the optic nerve, and divides into four flat muscle bellies that insert on the equator of the eyeball in a nearly complete muscular cone. Contraction of the m. retractor bulbi results in retraction of the eyeball into the orbit. Also, a series of four straight muscles originate at the apex of the orbit and project to the equator of the globe, superficial to the four bellies of the m. retractor bulbi. They are named for their insertion on the globe: mm. rectus dorsalis, rectus ventralis, rectus medialis, and rectus lateralis.
The m. obliquus dorsalis (dorsal oblique m.) lies between the dorsal and medial recti muscles and tapers to a thin tendon at the level of the posterior pole of the eyeball. This tendon passes around a small cartilaginous trochlea of cartilage anchored to the medial orbital wall. The trochlea redirects the pull of the tendon, which inserts on the dorsal part of the eyeball. When the dorsal oblique muscle contracts, the dorsal part of the eyeball is pulled toward the medial portion of the orbit (i.e., the left eyeball rotates counterclockwise and the right rotates clockwise).
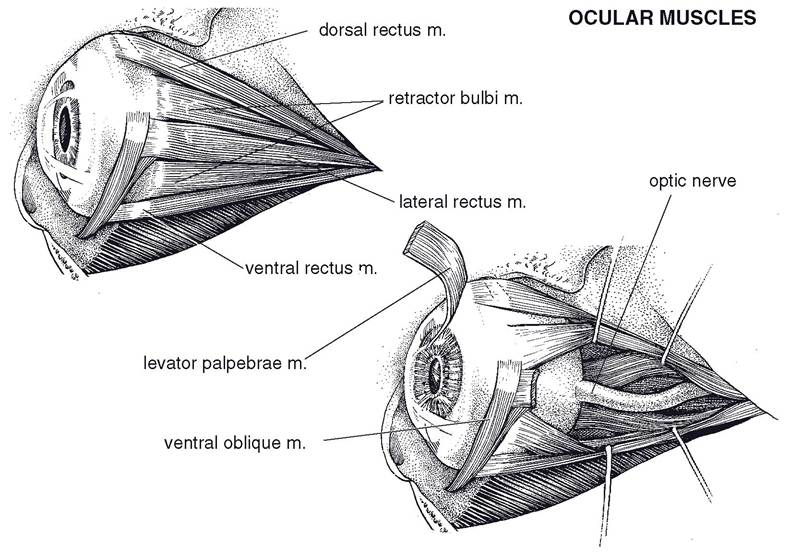
Figure 11-10. Extraocular muscles of the right eye viewed from the medial aspect.
The m. obliquus ventralis (ventral oblique m.) originates from a fossa in the ventral portion of the orbital rim; it runs dorsolaterad, ventral to the insertion of the ventral rectus muscle; and it inserts via two short tendons on the lateral aspect of the globe. The ventral oblique muscle rotates the globe opposite to the rotation produced by the dorsal oblique muscle.
One other muscle found within the orbit is not strictly an extraocular muscle, since it does not act on the eyeball itself. The m. levator palpebrae superioris is the primary lifter of the upper eyelid. It arises between the origins of the dorsal oblique and dorsal rectus muscles and inserts via a wide tendon in the connective tissue within the upper eyelid.
The periorbita is a cone-shaped connective tissue sheath that surrounds the eyeball and its muscles, nerves, and vessels. Like the extraocular muscles, the periorbita originates in the apex of the orbit; at the rim of the orbit, it blends with the periosteum of the facial bones.
The periorbita contains circular smooth muscle that squeezes its contents and places the eyeball forward in the orbit. Adipose tissue both within and outside the periorbita acts to cushion the orbital contents. Because of its location behind the eyeball, the adipose tissue is frequently called retrobulbar fat.
Globe
The eyeball (globe) comprises three concentric layers: the fibrous tunic, the vascular tunic, and the nervous tunic (Fig. 11-11). The three tunics of the eyeball surround several chambers filled with either liquid or a gelatinous material. The posterior 75% of the globe is filled with an acellular gel, the vitreous body, and as a consequence this portion is also called the vitreous chamber. The lens divides it from the anterior segment of the eyeball. The anterior segment of the eye is filled with a fluid called aqueous humor.
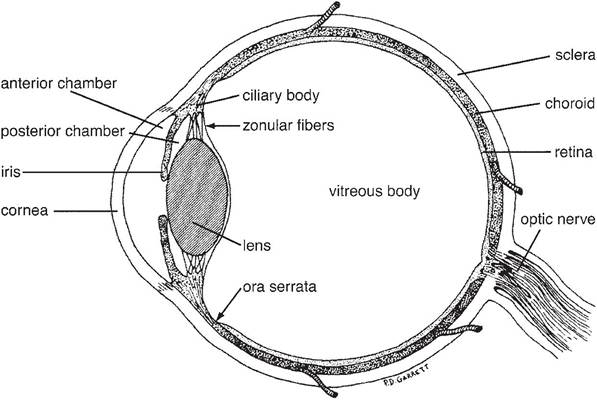
Figure 11-11. Sagittal section of a bovine eye. (Reprinted with permission of Wiley-Blackwell from Eurell, J.A. and Frappier, B.L. DellmannS Textbook of Veterinary Histology, 6th ed. Ames, IA: Blackwell Publishing Professional, 2006.)
Fibrous Tunic. The outer fibrous tunic of the eyeball is made up of a posterior opaque sclera and an anterior transparent cornea. Both are composed of very dense collagenous tissue. The sclera is white, variably tinged gray or blue; it meets the clear cornea at a transitional region called the limbus. The axons of the optic nerve pierce the sclera at the posterior pole of the eye, the area cribrosa. The tough sclera is the site of insertion for the extraocular eye muscles.
The cornea is the transparent anterior part of the fibrous tunic. It is the most powerful refracting layer of the eye (bends light more strongly than even the lens); its transparency and regular curvature are therefore critical elements for focusing of light on the retina. Corneal transparency is a function of (1) lack of vascular elements and cells, (2) lack of pigment, (3) relative dehydration of the collagenous tissue, (4) a smooth optical surface (provided in conjunction with the tear film), and (5) a highly regular, laminar pattern of collagen fibers that reduces light scatter. The cornea’s anterior and posterior surfaces are covered with specialized epithelium, but most of its thickness is composed of collagen fibers.
Vascular Tunic. The middle tunic of the eyeball, the vascular tunic, or uvea, is composed of three parts, the choroid, ciliary body, and iris. The uvea in the posterior portion of the eye is the choroid. It is highly vascular and possesses multiple layers. The deepest (closest to the center of the globe) of these is the tapetum. The tapetum of animals is a reflective surface that bounces incoming light back onto the retina and enhances vision in low light.
The shape and color of the tapetum are variable between species and individuals, but as a rule it is confined to the dorsal part of the posterior globe. The ventral portion of the choroid is usually not reflective. The pig and most primates lack a tapetum.The ciliary body is the anterior continuation of the uvea. It is a circumferential thickening of the vascular tunic, and it gives rise to many fine suspensory ligaments that support the lens. When the muscles in the ciliary muscle contract, they allow the lens to assume a more spherical shape; this increases its refractive power, a change that brings close objects into focus on the retina. This process of focusing on near objects is called accommodation. The capillaries of the ciliary body produce the aqueous humor in the anterior segment of the eyeball.
The iris is the most anterior portion of the uvea and the only part of the vascular tunic normally visible in the living animal. It consists of a pigmented ring of tissue, perforated in its center by the pupil. The iris divides the aqueous- filled anterior segment of the eye into anterior (in front of the iris) and posterior (between the iris and lens) chambers. The iris controls the amount of light entering the posterior part of the eye by changing the size of the pupil. Constrictor muscles, innervated by the parasympathetic fibers of the oculomotor nerve, reduce the size of the pupil when they contract. Dilation of the pupil is accomplished by activity of the dilator muscles, innervated by sympathetic nerves.
The iris derives its color from pigmented epithelial cells primarily on the posterior surface of the iris. The typical color distribution depends on the species and is related to the coat color of the individual. In herbivores, the pigmented epithelium of the posterior surface of the iris forms nodular masses along the margin of the pupil that are visible in the anterior chamber. These are often very large in horses, in which species they are called corpora nigra. In cattle the same structures are usually smaller and are called iridial granules. These appear to act as something of an “internal visor” for the pupil, shading it from direct sunlight.
The site where the anterior surface of the iris meets the fibrous tunic is the iridocorneal angle (fiItration angle); there the aqueous produced by the ciliary body is reabsorbed into the venous circulation. Abnormalities here (e.g., congenital narrowing or obstruction due to inflammation) can prevent normal egress of aqueous from the anterior chamber and result in increased intraocular pressure, a condition called glaucoma.
Nervous Tunic. The deepest layer of the eyeball is the internal (nervous) tunic or retina. The retina develops embryologically as an outgrowth of the diencephalon and is therefore composed of neurons and glia. The light-sensitive part of the retina extends from just posterior to the ciliary body to the site at which nerve fibers exit the eyeball near the posterior pole of the globe. In clinical practice the retina is routinely viewed through the pupil with an ophthalmoscope; the portion of the retina that can be viewed in this manner is called the fundus.
The retina has many layers. One of these is the photoreceptor layer, in which are found the specialized neural receptor cells of the visual system, the rods and cones. Each photoreceptor features an external segment that consists of orderly stacks of flattened, disk-shaped plasma membrane. These are replete with photopigment, the molecules that are sensitive to photons of light. Photoreceptors have one of two basic shapes: either a tall cylinder or a shorter, tapering stack. The cylindrical photoreceptors are rods, equipped with a photopigment that renders them sensitive to very low levels of light. The tapering photoreceptors are cones, each expressing one of three known photopigments with sensitivity to a specific wavelength of light. Cones are, therefore, color receptors. As a rule, animals with nocturnal habits have retinas populated primarily with rods, whereas those with diurnal habits (e.g., primates and birds) have more cones, particularly in the region that receives light from the center of the visual field.
Most domestic animals have some color vision. Their retinal cones are primarily sensitive to blue and yellow wavelengths of light, with few or no red-sensitive cones. Vision in these species is probably rich in blue, green, and yellow tones. The flapping red cape wielded by the matador therefore attracts the bull by its movement and not by its color.
The photoreceptors synapse with other neurons of the retina, and visual information undergoes early neural processing within the retina. Ultimately, the axons of neurons called ganglion cells project to the optic disk, where they exit the globe and become the optic nerve.
Lens. The lens is a transparent proteinaceous biconvex disk suspended between the posterior chamber and the vitreous chamber. It is surrounded by an elastic capsule that serves as an attachment site for the suspensory ligaments at the lens’s equator.
Cells of the lens are called lens fibers, and they are generated continuously throughout life. As these are added to the outside of the lens, older lens fibers are pushed toward the center, resulting in a lamellar arrangement of fibers, often likened to the layers of an onion.
As the animal ages, more lens fibers are added to the outside of the lens, compressing the fibers at the center. These old cells harden and after middle age begin to lose their transparency. This change is lenticular sclerosis. Although the sclerotic lens looks milky, lenticular sclerosis does not usually interfere with vision. More severe changes in the lens fibers can produce significant opacity that interferes with transmission of light and therefore vision. These opacities are cataracts. Cataracts can be congenital or can develop as a consequence of injury to the lens or metabolic disease (e.g., diabetes mellitus).
Visual Field and Light Path
The part of the environment from which light will enter the eyes and stimulate the retinas is the visual field (Fig. 11-12). In predators and arboreal animals, such as birds and primates (for whom accurate depth perception is essential), the eyes are placed so that the visual fields overlap to varying degrees. This region of overlap, where objects are simultaneously viewed by both eyes, is the binocular field; the visual cortex evaluates the slightly different view from each eye and uses the information to provide depth perception. Prey animals, on the other hand, have lateral eyes with a much smaller binocular field. such eye placement increases the peripheral vision so that the combined visual field is nearly completely panoramic. Such vision is monocular (seen only with one eye) and therefore lacks very accurate depth cues, but the clear advantage of this wide
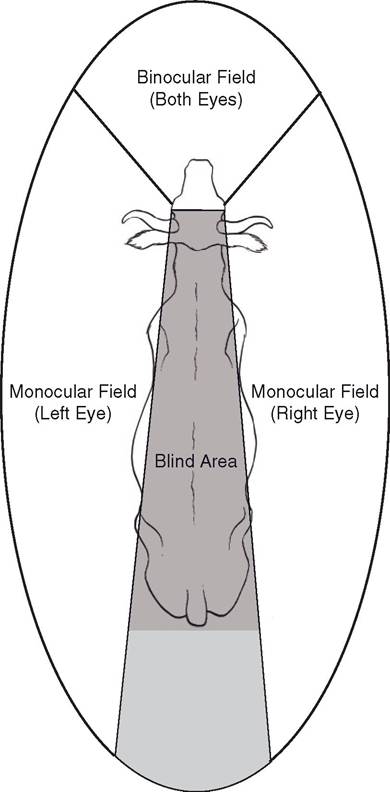
Figure 11-12. Visual fields of the ox as viewed from above. The region seen by both eyes (binocular field) is the region of best depth perception, but the nose creates a wedge-shaped blind spot directly in front of the animal. Peripheral vision (the monocular fields) creates a nearly complete circle of vision around the animal, except for about 3% directly behind.
field of view for a prey animal needs no explanation.
Light traveling from the visual field to the retina passes through a series of transparent media that refract and focus it on the lightsensitive retina of the posterior part of the globe. These dioptric media include the cornea, the aqueous, the lens, and the vitreous body. As indicated before, the cornea is actually the most refractive medium of the eye, but the lens is the only part of the light path with the ability to change its refractive index. This property makes it the organ of accommodation for focus on near objects. Light entering the vitreous chamber of the eye is bent by the more anterior parts of the eye in such a way that the image that is focused on the retina is inverted and reversed.
The site where the ganglion cell axons leave the eye (the optic disk) has no photoreceptors and is therefore considered the blind spot of the retina. In primates, a region dorsolateral to the optic disk is relatively free from large blood vessels and especially densely packed with photoreceptors (particularly cones). This is the region of greatest visual acuity, the macula. Domestic animals lack a macula.
Visual Pathways of the Brain
The electrical information generated by exposure of photoreceptors to light undergoes initial neural processing within the retina. This information ultimately leaves the eye via the optic nerve, the fibers of which are the axons of the ganglion cells of the retina. Most of the axons of the optic nerve synapse in the thalamus, and from there visual information travels to the primary visual cortex in the occipital lobe of the brain (the most caudal part of the cerebral cortex) for conscious perception.
A smaller subset of ganglion cell axons project to other destinations in the brain. Some reach the rostral colliculi of the mesencephalon, where visual stimuli induce reflex movements of the eyes and head. Others project to the pretectal nuclei, also in the region of the mesencephalon; these nuclei communicate with the oculomotor nuclei to coordinate the reflex constriction of the pupils in response to light. Finally, a very small number of ganglion cell axons project to a specific group of cells of the hypothalamus, the suprachiasmatic nucleus. The suprachiasmatic nucleus is the biologic clock, the part of the brain that sets circadian rhythms. Circadian rhythms are physiologic processes that vary regularly on a daily basis; prominent circadian rhythms include sleep-wake cycles, melatonin secretion, and body temperature fluctuations. The suprachiasmatic nucleus has an intrinsic rhyth- micity that closely approximates 24 hours, but the projections from the retina keeps the nucleus’s cycle entrained to the actual photoperiod of the day.
Many domestic species are seasonal breeders, meaning that their reproductive cycles are determined by the season. The most powerful determinant of the onset and cessation of breeding cycles in these species is the length of the day. The retinal projections to the suprachiasmatic nucleus are the brain’s record of day length, and they therefore determine the reproductive cycles via their influence on the autonomic functions of the hypothalamus. It is common agricultural practice to alter breeding behavior by exposing animals to artificial light. For instance, in the horse industry, in which an early foaling date is desirable, mares are commonly exposed to artificially increased day length in the winter so as to cause these spring breeders to begin fertile estrous cycles earlier than they would if exposed only to natural light.
There is a widely repeated myth among horse trainers that states that visual information from one side of the body is processed strictly on the opposite side of the brain and vice versa. Horse trainers have often cited this “fact” as a rationale for schooling horses from both sides of the body. While it is true that herbivores like horses process a majority of visual information from each half of their visual field in the contralateral visual cortex, there are three reasons why this idea is not neurobiologically sound. One is that not all of the visual information from each eye crosses to the contralateral cortex; some is processed on the ipsilateral (same) side.
Secondly, a small portion of the equine visual field is binocular, that is, is seen simultaneously by both eyes. And finally, the caudal part of the corpus callosum (the large bundle of axons connecting right and left cerebral hemispheres) connects the visual cortices of each side so that information is shared between the hemispheres. Trainers must look elsewhere for an explanation of why horses benefit from training on both sides.