Epidemiology of Paracoccidioidomycosis
PCM is restricted to Latin American countries (Figs. 6.1 and 6.2). In Brazil, which accounts for the majority of the cases, three large endemic areas are found: (1) Sao Paulo, Rio de Janeiro, Espirito Santo and Minas Gerais (southeast region), Goias and
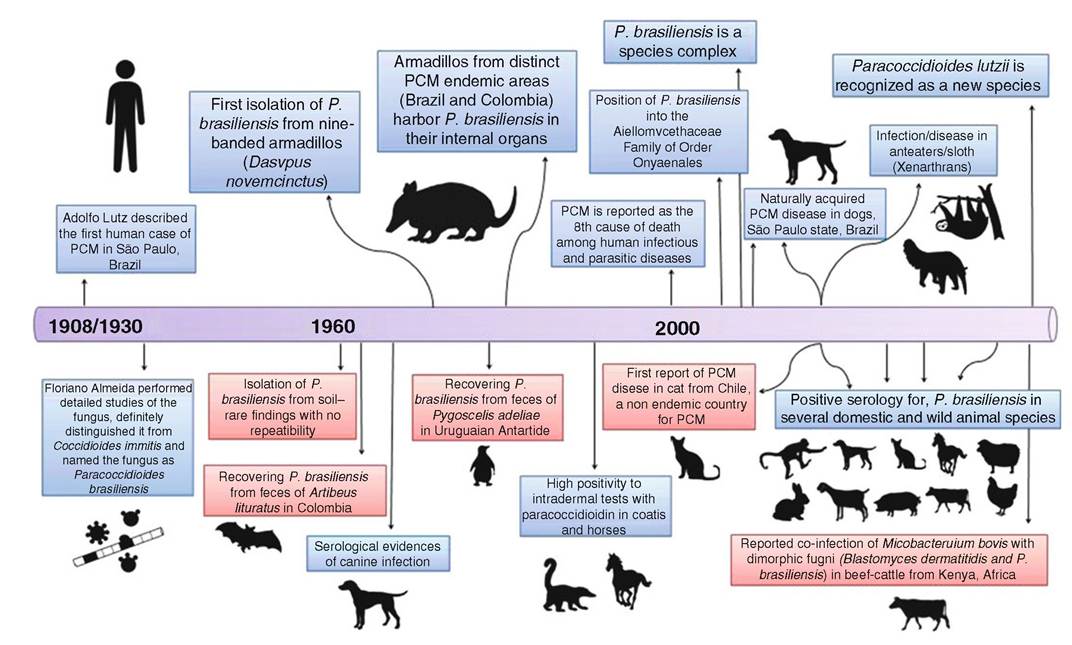
Fig.
6.2 The timeline of Paracoccidioides spp. and paracoccidioidomycosis eco-epidemiology. In red box, there are anecdotal findings with no repetition and/or confirmationMato Grosso do Sul (midwest region), and Parana to northern Rio Grande do Sul (southern region); (2) Para, Maranhao, and Tocantins (eastern of the Amazon region); and (3) Rondonia and the western Amazon region. Low endemicity of PCM is observed in the Brazilian northeast, a semiarid region where no autochthonous cases appear to exist in this semiarid region of Brazil. The endemic area of Mexico is located in its southern region, between the Gulf of Mexico and the Pacific coast. In Venezuela, areas endemic for PCM correspond to the northern region and in the state of Bolivar. Endemicity is also observed in Ecuador (Rio Cuenca Valley), in the region of the Peruvian Amazon forest, in the northeast and northwest of Argentina, in the eastern half of Paraguay, and in most of the Bolivian territory with a tropical climate. The PCM endemicity is low in Uruguay, while no autochthonous cases of the disease have been reported from Chile, Guyana, Surinam, French Guiana, Belize, or Nicaragua (Wanke and Londero 1994; Martinez 2015).
The ecology of Paracoccidioides species is still poorly known, and the fungus continues to challenge researchers in relation to its ecological niche (Restrepo 1985; Restrepo-Moreno 1994; Restrepo et al. 2001; Bagagli et al. 2012). While it is known that the fungus exists in nature as a saprobe in soil, it has only rarely been cultured from soil or soil-related products, including feces of both frugivorous bats (Artibeus Iituratus), penguins (Pygoscelis adeliae), and dog food contaminated with soil (Franco et al.
2000; Restrepo et al. 2000). These reports were difficult to reproduce and are not conclusive with respect to transmission of the fungus (Bagagli et al. 2012).Experimental PCM infection has been well studied and documented in several animal models, such as in mice, rats, hamsters, and guinea pigs (McEwen et al. 1987; Tani et al. 1987; Singer-Vermes et al. 1993; Coelho et al. 1994), as well as in dogs and cattle (Mos et al. 1974; Costa et al. 1978; Ono et al. 2003; Eisele et al. 2004). Natural PCM infection in domestic and wild animals has been detected by intradermal tests, serological surveys, histopathological analysis, molecular biology, and by isolation of the pathogen in culture. The first studies in dogs that employed complement fixation and precipitation tests revealed seropositivity in almost 80% of the animals evaluated, which originated from recognized human endemic areas of PCM (Mos and Netto 1974; Mos et al. 1974). Intradermal tests using paracoccidioidin as antigen among domestic animals, cattle, horses, and sheep, yielded positive results in 40.2, 63.8, and 40.8%, respectively (Costa et al. 1978, 1995a; Costa and Fava Netto 1978). Intradermal tests in terrestrial wild animals (coatimundis and Felidae) present higher positivity than in arboreal animals (weeping capuchins and marmosets), i.e., 82.98 and 22.45%, respectively (Costa et al. 1995b).
Additional epidemiological studies using sensitive serological approaches, such as ELISA, Western blotting, and immunodiffusion, as well as histopathology and nested PCR, have confirmed that natural PCM infection occurs frequently in several domestic and wild animals from endemic areas. As to dogs, high positivity was confirmed among animals from PCM-endemic areas of Brazil (Silveira et al. 2006; Teles et al. 2015) and Argentina (Canteros et al. 2010), especially when in rural areas, where positivity may reach 80.5-89.5% (Ono et al. 2001; Fontana et al. 2010). It is important to emphasize that serological surveys were carried out on puppies and young and adult dogs of both sexes and of different breeds.
No significantly different prevalence was observed with sex, breed, or age; the origin of the dog (rural vs. urban) was the most important parameter. PCM infection was also observed in cats (de Oliveira et al. 2013), chickens (Oliveira et al. 2011), dairy cattle (Silveira et al. 2008), pigs (Belitardo et al. 2014a), dairy goats (Ferreira et al. 2013), rabbits (Belitardo et al. 2014b), and sheep (Oliveira et al. 2012), as well as in wild mammals, such as the capuchin monkeys (Cebus sp.) and golden howler monkeys (Alouatta caraya) (Corte et al. 2007). Infections were also noted in the small rodents (Akodon sp., Thaptomys nigrita, Euryoryzomys russatus, and Oligoryzomys nigripes) (Sbeghen et al. 2015), road-killed nine-banded armadillos (D. novemcinctus), seven-banded armadillos (D. septemcinctus), guinea pigs (Cavia aperea), porcupines (Sphiggurus spinosus), grisons (Galictis vittata), and raccoons (Procyon cancrivorus) (Richini-Pereira et al. 2008b).The nine-banded armadillos and the naked-tailed armadillo (Cabassous centralis) carry P. brasiliensis in their internal organs (Naiff et al. 1986; Bagagli et al. 1998, 2003; Corredor et al. 1999, 2005; Silva-Vergara et al. 2000; Richini-Pereira et al. 2008a). In some endemic areas, all evaluated armadillos provided positive culture, especially from fragments of mesenteric lymph nodes and to a less extent from spleen and liver tissues (Bagagli et al. 2003) (Fig. 6.3). There is no clear evidence as to whether infected armadillos develop signs of active PCM disease, despite few granulomas containing fungal elements observed in lymph nodes, liver, and lungs of some animals (Bagagli and Bosco 2008). Armadillos originated in South America in the same geographic area as PCM. Besides intense contact with soil where they live as terrestrial animals, armadillos present a low body temperature and a weak cell- mediated immunity, which render them particularly susceptible to fungal infection (Bagagli and Bosco 2008). Other xenarthran species, such as the seven-banded armadillo, anteater, and sloth, are also infected, being positive by nested PCR with primers specific for P. brasiliensis or histopathology (Richini-Pereira et al. 2008b, 2009, Trejo-Chavez et al. 2011).
6.4
More on the topic Epidemiology of Paracoccidioidomycosis:
- Epidemiology of Paracoccidioidomycosis
- References
- Seyedmousavi S. et al. (eds). Emerging and Epizootic Fungal Infections in Animals. Springer International Publishing,2018. - 406 p, 2018
- Abstract
- Index
- References