Equine Respiratory Viruses
Gabriele A. Landolt
Viral respiratory infections in horses are major causes of respiratory disease resulting in impaired health, poor performance, and financial losses. Although vaccines are available for many of the etiologic agents, viral respiratory disease has remained among the most commonly encountered equine health concerns worldwide.
Infection with equine respiratory viruses can lead to serious respiratory illness with potentially fatal consequences, yet the clinical significance of some of the viruses that have been detected in equine respiratory tract secretions remains largely unknown (e.g., EHV-2, EHV-5, equine coronavirus).1 The severity of equine viral respiratory disease depends on the agent, virulence of the circulating virus, infectious dose, management and environmental conditions, and degree of immunity of the individual animal. In a group of susceptible horses, equine viral respiratory disease is often characterized by the rapid spread of an acute, febrile respiratory disease that may be accompanied by a cough and serous nasal discharge. Therefore a presumptive diagnosis of viral respiratory disease can often be made based on history and clinical signs. However, to confirm and differentiate among the potential etiologic agents, laboratory diagnosis is required.
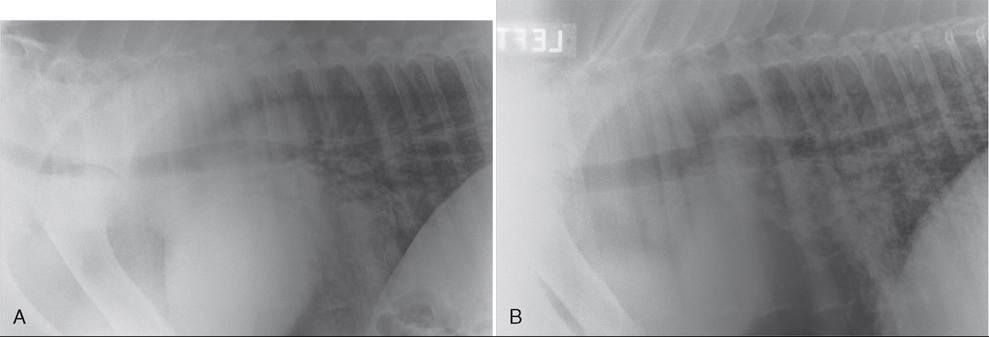
FIG. 31.31 A, Midthoracic/caudal dorsal thoracic radiograph of a foal younger than 2 months that presented with Pneumocystis spp. pneumonia and acute respiratory distress syndrome (ARDS). Immune function testing was within normal limits. Note the predominant interstitial pattern with patchy alveolar component. B, Similar radiographic view of the foal in A following 4 weeks of treatment with antimicrobials and corticosteroids. This foal was also treated with inhaled antimicrobials, inhaled corticosteroids, and intranasal oxygen insufflation over the first 2 weeks of treatment.
The foal was significantly improved clinically and radiographically.■ Diagnosis of Equine Respiratory Viruses The basic approaches for the laboratory diagnosis of equine viral respiratory disease are the isolation of the etiologic agent (virus isolation); demonstration of the virion, the virus genome, or the viral products in clinical specimens (detection of virus particles by electron microscopy; detection of viral antigens by immunofluorescence, ELISA, or immunoperoxidase; detection of the viral genome); and the detection of virus-specific antibodies (serology). Because each of these methods has merits as well as disadvantages (e.g., lack of sensitivity, long turnaround time, high costs), it may be necessary to combine several of them to identify the etiologic agent accurately and rapidly. The World Organisation for Animal Health (OIE; http://www.oie.int) publishes the Manual of Diagnostic Tests and Vaccinesfor Terrestrial Animals, which provides an in-depth review of current diagnostic testing strategies.
VIRUS ISOLATION. Virus isolation from clinical samples is crucial for epidemiologic investigation and for vaccine production and generally is carried out in cell culture. The notable exception is equine influenza virus, for which embryonated chicken eggs have remained the culture system of choice. Because virus shedding in secretions of the respiratory tract is typically greatest within the first few days after infection, the best results for virus isolation are often achieved by culturing nasal or nasopharyngeal swab samples collected in the first 24 to 48 hours after onset of clinical disease. Because the duration of nasal virus shedding is often brief in partially immune animals, it may prove useful to sample immunologically less protected individuals in a group (e.g., younger horses, poorly vaccinated or unvaccinated animals) to increase the likelihood of demonstrating infectious virus. Nasopharyngeal or nasal swab samples are best collected using polyester-tipped swabs instead of cotton swabs because certain viruses (e.g., influenza A virus) have been found to adhere to the cotton fibers.
Preferably, the swab samples should be placed in a small amount of sterile viral transport medium and kept on ice.ANTIGEN DETECTION. The most important advantages of diagnostic tests aimed at the detection of viral antigen over traditional virus isolation are the faster turnaround time as well as the ability to detect virions that have lost their infectivity. Immunofluorescence (fluorescent antibody [FA] test) is based on the detection of virus-infected cells using virus-specific fluorochrome-labeled antibodies. The technique is used to detect viral antigen in a broad range of clinical samples (e.g., frozen sections of tissues, tissue imprints, cells obtained from nasal scrapings or tracheal washes) and has been shown to be highly sensitive. A number of antigen-capture ELISA-based assays have been developed for the detection of equine respiratory viruses.2-6 Frequently, a solid-phase system is used for virus detection. Specific antibody is first adsorbed to the surface of a microtiter plate, and the sample containing the suspected virus is then added. If the virus is present, it binds to the adsorbed antibody. After washing, specific enzyme-labeled antibodies are added. Detection of virus in the sample is achieved by addition of the enzyme substrate, which results in a color reaction. Commercially available ELISA-based test kits (e.g., Flu OIA® assay [Biostar, Boulder, Colo.]; Directigen Flu-A assay® [Becton Dickinson Microbiology Systems, Cockeysville, Md.]) have been validated for the rapid, stall-side detection of equine influenza virus. Many investigators found these commercial diagnostic kits to be highly specific and rapid, and their sensitivity approaches that of egg-based virus isolation.7-11 Since the discovery of PCR, the technique has found wide application in diagnostic laboratories. By choosing appropriate oligonucleotide primers, a selected region of the viral genome can be amplified. PCR-based assays are extremely sensitive and theoretically can detect a single copy of the target nucleic acid in a sample.12-14 Due to the assay's high sensitivity, the greatest challenge facing the diagnostic application of PCR is the production of false-positive results.
Nevertheless, PCR-based techniques offer a more sensitive tool for the diagnosis of equine respiratory viruses than conventional techniques such as culture or immunoassays. To increase the diagnostic capacity of PCR, multiplex PCR assays have been developed. By including more than one pair of primers in the reaction mixture, these assays are designed to simultaneously detect a panel of respiratory pathogens. RT-PCR-based assays are widely used for the detection and quantification of equine respiratory pathogens.15-17 By using a target-specific fluorescent probe, RT-PCR-based assays eliminate the need to distinguish the target by PCR fragment size on electrophoresis or hybridization with probes post PCR. The advantages of RT-PCR include high specificity, sensitivity, and a rapid turnaround time.ANTIBODY DETECTION. Detection of virus-specific antibodies has been and continues to be a key tool in the diagnosis of equine viral respiratory disease. Detection of specific antibody can provide support that an animal has been exposed to a given antigen. Although this is often used as circumstantial evidence of infectious disease, a diagnosis of active infection requires the demonstration of at least a fourfold increase in antibody titers (seroconversion) in samples collected during acute infection and during convalescence. Therefore serology typically provides only retrospective information. Yet as most serologic assays are fairly easy to perform and cost-effective and because large numbers of samples can be collected and tested simultaneously, serologic testing is particularly useful for large-scale population surveillance. Commonly used techniques for antibody detection include ELISA-based assays, virus neutralization tests, single radial hemolysis, complement fixation tests, and hemagglutination inhibition assays. Due to the inherent variability of immunoassays, paired samples always should be run by the same laboratory. Lastly, it is important to recognize that antibody is not always detected in the presence of active infection.
The most common reason for a negative result despite infection is that insufficient time has passed between infection and measurement of an antibody response. Alternatively, immunocompromised horses may fail to produce a detectable antibody response despite active infection.■ Treatment The treatment of horses suffering from viral respiratory disease is primarily symptomatic. It is important to ensure that sick animals are kept in a clean, well-ventilated, and stress-free environment. Morbidity caused by pyrexia and myalgia may be reduced by administering NSAIDs. It is important to monitor affected horses for development of complications such as pneumonia and myocarditis, and animals exhibiting signs of respiratory disease beyond 10 days should be reevaluated to rule out secondary bacterial infections.
Antiviral drugs, such as amantadine, rimantadine, oseltamivir, and the chain-terminating nucleoside analogs (e.g., acyclovir, valacyclovir, cidofovir), have been evaluated for prophylaxis and treatment of horses during outbreaks of equine influenza and EHV-1, respectively. In addition, there are several published reports describing the use of acyclovir in horses diagnosed with EMPF.18,19
Clinical trials with the influenza virus M2 ion channel blockers amantadine and rimantadine have shown that both drugs appear to be equally effective in reducing the severity and duration of clinical illness.20,21 In addition, oral administration of rimantadine (30 mg/kg q12h) was shown to reduce the virus load in nasal secretion, although the duration of nasal virus shedding was similar to that in the untreated controls.20 However, the therapeutic and prophylactic benefits of M2 ion channel blocker antiviral therapy during outbreaks of equine influenza may be limited due to the potential for the rapid development of drug resistance. In vivo and in vitro studies have demonstrated that drug-resistant influenza virus mutants can develop as early as 1 day after start of treatment.22 Subsequent isolation of the drug-resistant virus from susceptible contacts showing signs of clinical disease indicates that acquisition of drug resistance is not associated with attenuation of the virus.23,24 Treatment of horses with amantadine has also been associated with adverse effects and poor oral bioavail- ablity.25 Oseltamivir, a neuraminidase inhibitor with demonstrated antiviral activity against equine influenza in vitro, was also found to reduce the magnitude of nasal virus shedding and severity of clinical signs in horses following experimental inoculation.26 In addition, oseltamivir administration reduced the counts of S.
equi subsp. Zooepidemicus in bronchoalveolar fluid samples collected 7 days after influenza infection compared to the untreated controls.26 Despite these findings, the use of antivirals during an equine influenza outbreak should be carefully deliberated, as their use can result in the development of antiviral resistance among influenza A viruses.27-29 Hence the indiscriminate use of these drugs can possibly reduce the effectiveness of treatment during an outbreak.Acyclovir, valacyclovir, ganciclovir, valganciclovir, and other chain-terminating nucleoside analogs have been shown to be efficacious against various strains of abortigenic and neuro- pathogenic EHV-1 viruses in vitro.30-33 Despite the fact that the oral bioavailability of acyclovir in horses is low,34 anecdotal reports suggest a potential benefit of acyclovir administration (10 mg/kg PO five times daily) in horses with clinical signs of EHV-1 infection (e.g., herpes myeloencephalopathy and neonatal infection).35-37 Studies examining the effectiveness of oral valacyclovir, a prodrug of acyclovir with enhanced oral absorption, in the treatment of horses infected with EHV-1 have demonstrated controversial results. Although administration of valacyclovir (27 mg/kg q8h followed by 18 mg/kg q12h) to aged mares either prior to experimental EHV-1 inoculation or at the onset of fever reduced the severity of clinical signs, virus shedding, and viremia in one study,38 another study found no effect on clinical signs, nasal virus shedding, or viremia in ponies treated (40 mg/kg q8h) compared to the untreated control ponies.39
Equine Influenza Virus
Equine influenza is the most frequently diagnosed cause of viral respiratory disease of the horse.40,41 It is a highly infectious disease that affects horses, donkeys, mules, and zebras. Outbreaks of a disease resembling influenza have been reported as early as 1751; however, the etiologic agent was not isolated until 1956.42,43
■ Etiology Equine influenza viruses are members of the family Orthomyxoviridae, which contains enveloped viruses with segmented, single-stranded, negative-sense RNA genomes.44 The Orthomyxoviridae comprise the following genera: influenza A, B, and C viruses; thogotoviruses; and isaviruses.44 Influenza D virus was isolated from pigs and cattle,45,46 and serologic evidence for influenza D virus infection in humans and horses has recently been established.47,48 Despite this, the clinical significance of this finding is currently unknown.
Equine influenza viruses are members of influenza A viruses. In contrast to influenza B and C viruses, which appear to have a fairly narrow host range, influenza A viruses can infect a wide variety of species (e.g., horses, humans, wild and domestic birds, pigs, sea mammals, dogs, and cats). It has long been recognized that influenza A viruses exhibit partial restriction of their host range,43 indicating that viruses from one host species occasionally can transmit to infect another host. Although horses have often been regarded as isolated or “dead-end” hosts for influenza,43 events of recent years clearly indicate that the species barrier for viruses jumping either to or from horses is not absolute. For example, the severe influenza epidemic in horses occurring in 1989 in the Jilin and Heilongjiang provinces in the northeast of the People's Republic of China was caused by a virus with avian-like genetic features, suggesting that the virus had spread directly to horses from birds.49 Another, more recent example is the transmission of an equine-lineage H3N8 virus to dogs in the United States.50
Although the viral and host factors that determine the host range of influenza viruses and the mechanisms by which species barriers are overcome are not completely understood, a large body of research highlights the importance of the viral hemagglutinin (HA) glycoprotein in determining host specificity.43,51,52 The HA serves as the viral receptor-binding protein and is embedded, along with the neuraminidase (NA) and the ion channel M2 protein, in a host cell-derived lipid envelope. Mediated by binding of the HA to sialic acid residues on the host cells, the virion envelope and the host cell membrane fuse. This interaction subsequently promotes the release of the viral genome into the cytoplasm of the host cell. The NA is responsible for the cleavage of the α-ketosidic linkage between a sialic acid molecule and an adjacent sugar D-galactose or D-galactosamine. Biologically, the NA facilitates the mobility of the influenza virus virion by assisting in the release of the budding virus particle from the host cells.53,54 Both HA and NA are major surface antigens of the influenza virus virion. Partly due to the host immune pressure, these proteins can undergo substantial antigenic variation (antigenic drift), which allows the mutant viruses to escape neutralizing antibodies.
Based on antigenic properties of the HA and NA, influenza A viruses are divided into subtypes. To date, eighteen HA subtypes (H1 to H18) and eleven NA subtypes (N1 to N11) have been described.55,56 With the exception of two HA and NA subtypes (H17, H18, N10, N11) that have been isolated only from bats, all other subtypes have been isolated from wild waterfowl.55,57-59 In contrast, only a limited number of subtypes have been associated with infection of mammals. In humans, only viruses of H1, H2, H3, N1, and N2 subtypes have circulated widely in the population,43,60 and only H1, H3, N1, and N2 subtypes have been consistently isolated from pigs.61-63 In horses, two influenza subtypes have been described: H7N7 (A/equine/1) and H3N8 (A/equine/2).42,64,65 Although there are a few reports of horses with serologic evidence of H7N7 exposure,42,66,67 the H7N7 viruses have not been isolated from horses since the late 1970s11,57,68 and are considered to be extinct. In contrast, despite intensive vaccination programs, equine H3N8 viruses continue to circulate in large parts of the world. Since the 1980s, a number of severe, widespread influenza outbreaks in horses have occurred in South Africa,69 India,70 the People's Republic of China,49,71 the United Kingdom,72 and most recently South America.73,74 In 2007 an equine influenza outbreak occurred for the first time in Australia, a country previously considered to be free of the disease.75 The outbreak was caused by an equine-lineage H3N8 virus that was thought to have been introduced by importation of influenza-infected horses from Japan.75,76 Stringent control measures, consisting of movement restrictions and vaccinations, allowed for the eradication of the virus from the Australian
horse population, and Australia regained its equine influenza- Jr Jr ^ D TL
free status in December 2008.77,78
Since the early to mid-1980s, the equine H3N8 viruses have diverged into two distinct evolutionary lineages: Eurasian and American. Although the circulation of both lineages initially centered largely on their geographic origin,79 introduction and apparent maintenance of American lineage viruses in horses in Europe initially resulted in the co-circulation of both lineages in Europe. In contrast, only a few reports support sporadic infection of horses in North America with viruses originating in Europe.80-82 Continued genetic divergence of the American lineage viruses resulted in the formation of three antigenically distinct American-like sublineages (a South American lineage, a Kentucky lineage, and a Florida lineage).82 Since the early 2000s the Florida sub lineage has further evolved into two antigenically distinct groups, referred to as Florida sublineage clades 1 and 2. Whereas clade 1 viruses primarily circulate in North America, clade 2 viruses have mainly been identified in Europe and parts of Asia.80,82 Despite the fact that the rate of genetic diversion of equine viruses is small compared to human and swine influenza viruses, this sustained genetic and antigenic evolution continues to have an impact in terms of immunization.
■ Epidemiology Experimentally, horses of all age-groups are susceptible to infection.83 However, although sporadic infection in foals has been described, there exists only one published report of a widespread influenza outbreak in foals younger than 6 months of age.84 The lower incidence of disease in young foals is most likely due to the presence of maternally derived antibodies. Influenza-specific serum antibody concentration (particularly antibody titers measured by single radial hemolysis) is often used as a correlate for protection against infection and disease, and animals with high concentrations of homologous antibody are by and large protected against experimental challenge.85,86 After natural infection, ponies were reported to be protected from infection for 8 months, with partial protection persisting for more than 1 year.87 Although vaccination does not always provide full protection against clinical disease, immunization has reduced the frequency of influenza outbreaks and the frequency and severity of clinical 838889 signs when influenza occurs among vaccinated animals.83,88,89
Equine influenza outbreaks occur most often when susceptible animals are congregated and housed in close contact with each other (e.g., horse shows, racetracks, sale barns). It is thought that spread of virus among susceptible animals occurs through three modes: direct contact with infected animals or fomites, droplet transmission (droplets larger than 10 μm and capable of being projected over moderate distances by coughing and sneezing), and aerosol transmission (droplet nuclei smaller than 5 μm, capable of wide dissemination and of reaching the lower respiratory tract).90 Although aerosol transmission clearly has the greatest impact in regard to influenza virus infection control, the importance of bioaerosol transmission in the spread of influenza virus is poorly understood. Depending on the prevailing environmental conditions (humidity, temperature, exposure to sunlight, etc.), the virus can remain infectious for days on contaminated surfaces.91 Meteorologic factors associated with an increased risk of spread of equine influenza virus between premises during a recent outbreak were relative humidity of less than 60% and wind speeds greater than 30 km/h from the direction of a barn with infected horses.92
Once the virus has been introduced into a group of immunologically naive animals, disease can spread rapidly (in a matter of hours to days). Such effective spread is enhanced by a short (~2 days) incubation period, high concentrations of virus shed in nasal secretions, a forceful cough, as well as the practice of housing horses in confined spaces.91 In partially immune animals, the spread of disease is often considerably slower, and outbreaks may last as long as 3 to 4 weeks.93 Following experimental infection, horses typically shed virus for 6 to 7 days.94-97
■ Pothogenesis Influenza A viruses replicate and induce pathologic changes throughout the respiratory tract, with the most significant lesions typically occurring in the lower airways. After inhalation of the virus, the virion attaches to sialic acid receptors on the surface of the respiratory epithelial cell and enters the cell by receptor-mediated endocytosis. Following replication and release from the host cell, the virus spreads quickly throughout the respiratory tract. Largely through
98100
apoptosis,98-100 influenza virus replication leads to cell death. The loss of the ciliated respiratory epithelium in the trachea and bronchial tree101 results in the impairment of tracheal mucociliary clearance.64,102,103 In addition, the disruption of the superficial layers of the respiratory epithelium allows opportunistic bacteria to invade the deeper tissues of both the upper and the lower respiratory tract. The combination of reduced airway clearance and erosion of the respiratory epithelium predisposes affected horses to the development of secondary bacterial complications such as bacterial bronchopneumonia.101,104 In uncomplicated cases, regeneration of the respiratory epithelium begins about 3 to 5 days after onset of illness,104 yet resolution of the epithelial damage takes a minimum of 3 weeks.103,105
■ Clinical Presentation Clinical signs of influenza virus infection in horses are often seen 48 hours after exposure to the virus. In vaccinated animals, the disease is rarely fatal, but deaths have been reported during some epidemics, particularly in donkeys and rarely in neonatal foals.42,106 The typical clinical presentation of equine influenza consists of pyrexia, anorexia, lethargy, nasal discharge, and cough. Pyrexia is often the first symptom present, with body temperatures peaking somewhere between 48 to 96 hours after infection. In some cases, a second peak of pyrexia may occur around 7 days after infection. Nasal discharge is typically serous in the first few days of illness but may become mucopurulent by 72 to 96 hours after infection. Concurrently, a dry, hacking cough develops. Coughing tends to persist long after pyrexia and nasal discharge have resolved and may still be present 3 weeks post infection. The majority of affected animals are anorexic during the initial pyrexia. Uncomplicated cases of equine influenza can resolve within 7 to 14 days post infection. Complications of equine influenza can be severe and include secondary bacterial pneumonia, myositis, myocarditis, and limb edema.65,102,103 Neurologic disease as a result of a nonsuppurative encephalitis has also been described.107 Finally, it has been hypothesized that influenza infection may predispose horses to the development of equine asthma and exercise-induced pulmonary hemorrhage.65,101,102 The effects of influenza virus infection can be significantly exacerbated by even moderate exercise.108
Hemogram abnormalities associated with equine influenza infection can include a moderate normocytic, normochromic anemia and a leukopenia, which may result from both a neutropenia and lymphopenia. Monocytosis during early convalescences is a variable finding.
■ Control Control of equine influenza primarily is based on adequate husbandry procedures and vaccination. Introduction of equine influenza virus into a group of horses likely can be prevented by following the biosecurity guidelines (e.g., isolation of horses for 4 weeks prior to introduction into the horse population) published by the OIE (published in the Terrestrial Animal Health Code, Volume II, 2013; http://www.oie.int/). Because these biosecurity measures may be difficult to implement for most barns, 2 weeks of strict isolation and adequate vaccination may serve as an adequate compromise. Unfortunately, even these measures are often not taken, and vaccination is the primary modality used for infection control.
Although a wide variety of vaccine formulations, including modified live and recombinant vaccines, are commercially available, inactivated vaccines remain the most common type of vaccine used. Despite the fact that the vaccines are generally very efficacious, outbreaks of equine influenza continue to occur. One of the reasons for this is that the virus undergoes antigenic drift, which results from the accumulation of amino acid changes in the antigenic sites of the hemagglutinin protein. Studies have shown that even a single amino acid difference in one of the five antigenic sites of the hemagglutinin protein between the vaccine and infecting virus strain can result in a reduction of the protection induced by the vaccine.109,110 In addition, vaccination seldom induces the same immunity as natural infection, and vaccine-induced antibodies may not persist for very long.100 This is particularly true for vaccines that are based on inactivated whole virus. In contrast, vaccines using modified live virus and recombinant vectored virus technologies (e.g., canary poxvirus-based vaccines) stimulate both humoral and cellular immune responses and mimic immunity elicited by natural infection more closely.100
Similar to human influenza virus vaccines, equine vaccines need to be updated periodically to include relevant contemporary virus strains. Vaccine strain recommendations are based on epidemiologic, genetic, and antigenic data that are analyzed by a panel of equine influenza specialists overseen by the OIE.111 With the divergence of the Florida sublineage, current recommendations are that killed vaccines should include strains representative of both Florida clade 1 and clade 2.112 Because the sustained genetic evolution of the equine H3N8 viruses will continue to result in vaccine failures in the future, inclusion of viral antigens representative of contemporary circulating viruses must remain a priority.
Studies have demonstrated that an intranasal, cold-adapted, modified live virus vaccine was able to confer protection for up to 12 months after a single administration. Despite this, only a 6-month claim is made on the product data sheet.97,113 While the vaccine contains only one modified life virus strain, which is based on the H3N8 A/Eq/KY/91 isolate, the vaccine appears to protect against both European and American lineage isolates.114 Lastly, a recombinant canary poxvirus vector-based vaccine has also shown excellent performance against recent influenza virus strains.115
Recommendations for equine vaccination are available from a variety of sources. Due to the presence of maternally derived antibodies, foals should not be vaccinated before at least 6 months of age.116-118 For inactivated vaccines, initial vaccination series should include three doses even when the data sheets recommend only two initial doses.117 An interval of 3 to 4 weeks between the first and second doses is recommended, but a longer interval of 3 to 4 months between the second and third doses is preferred. This results in the third dose of the initial series being administered when the antibody response to the second vaccine dose has waned, and the amplitude of the antibody response to the third dose is consequently much greater.119 For high-risk populations (show horses, racehorses, etc.), booster vaccinations should be given at 6-month intervals.117,120 Additional booster doses may be administered 1 to 2 weeks prior to potential exposure if a higher risk of infection is anticipated. The modified live intranasal vaccine requires only a single initial dose, followed by booster doses at 6-month intervals.
Equine Herpesviruses
Equine herpesvirus infection is a common infection of equids throughout the world, with at least nine herpesviruses identified to date.121,122 The equine herpesviruses with the most clinical importance are the Alpbaherpesvirinae EHV-1 and EHV-4, which both cause respiratory disease. EHV-1 commonly infects horses, donkeys, and other equids and occasionally infects domestic cattle, captive camelids, cervids, and zebras; EHV-4 is restricted to horses.121 EHV-3, also a member of the subfamily Alphaherpesvirinae, is the cause of equine coital exanthema, a venereally transmitted disease characterized by superficial lesions on the epithelium of the external genitalia of mares and stallions.
Members of the Gammaherpesvirinae subfamily, both EHV-2 and EHV-5 have been associated with a range of clinical diseases, including respiratory disease. Whereas EHV-2 has been associated with poor athletic performance, respiratory disease, and keratoconjunctivitis,123-126 EHV-5 has been hypothesized to be the causative agent of EMPF and equine lymphoma.127-129 EMPF is a progressive pulmonary disease characterized by coalescing nodules of fibrosis and interstitial pulmonary fibrosis and pneumonia.127 Results from a recent study in which EMPF was experimentally induced after inoculation of horses with virus isolated from EMPF lesions provides added support for the causal relationship between EHV-5 infection and the disease.130
Studies aimed at determining the clinical significance of EHV-2 and EHV-5 infection are complicated by the ubiquitous nature of infection in the equine population. EHV-2 and EHV-5 have been detected in horses throughout the world.131-133 As both viruses can frequently be detected in samples obtained from healthy horses as well as from horses with clinical disease, the relevance of the finding remains unknown.134-137
■ Etiology Both EHV-1 and EHV-4 are members of the Alphaherpesvirinae subfamily of herpesviruses, in the genus Varicellovirus. These two viruses have very similar antigenic and genetic characteristics, although they are quite distinct from the gammaherpesviruses EHV-2 and EHV-5.121 Both viruses are easily inactivated by solvents, detergents, disinfectants, and other common veterinary means of decontamination but in favorable environmental conditions can persist for as long as a month and in refrigerated liquids for much longer.
Both EHV-1 and EHV-4 have a linear double-stranded DNA genome of 145 and 150 kbp encoding 79 and 80 open reading frames (ORFs), respectively,138-140 which can code for 77 proteins. Amino acid homology of EHV-1 and EHV-4 is 55% to 96%.138 The gene products of EHV-1 and EHV-4 are classified as immediate early, early, or late (which include the major glycoproteins).141-144 In recent years, a single nucleotide polymorphism (N752/D752) in the EHV-1 ORF 30, encoding the catalytic subunit of the viral DNA polymerase, was suggested to be significantly associated with the neuropathogenic potential of EHV-1 strains.145,146 In support of the increased neuropathogenic potential of the N to D substitution at position 752 are the results of a recent study using recombinant viruses with either genotype.145 Although results of several recent studies indicate that the less virulent genotype (N752) appears to be 147149
more prevalent in the equine population at this time,14' 149 viruses with D752 have increased in prevalence in recent decades.150 It has been hypothesized that the link between the substitution of asparagine (N) and aspartic acid (D) at position 752 and its neuropathogenic potential is due to the higher magnitude and duration of viremia that was found after experimental infection of horses with virus carrying this mutation.151 In contrast, a recent study found no significant difference in viral loads in nasal secretion and blood between the two genotypes in horses with natural EHV-1 infection.149 The discovery of this mutation and its association with neuropathogenic potential led to the development of PCR-based diagnostic assays that are able to detect strains carrying the N752D mutation. Despite this, it should be taken into consideration that infection with an EHV-1 strain encoding D at position 752 does not inevitably result in neurologic disease. Likewise, although D752 is present in the large majority of EHV-1 viruses causing neurologic disease, equine herpesvirus myeloencephalopathy (EHM) has also been caused by strains with N at position 752. Lastly, glycoprotein D, an essential protein in the viral entry process, has recently been shown to be another important virulence factor of EHV-1 in horses.152
■ Epidemiology EHV-1 infection causes significant suffering in horses and extensive economic losses through frequent outbreaks of a range of diseases, including respiratory disease, abortion, neonatal foal death, EHM, and chorioretinopathy.121 EHV-4 can cause respiratory disease outbreaks but only sporadic cases of abortion rather than abortion storms. Infections caused by EHV-1 and EHV-4 are particularly common in young performance horses and frequently result in establishment of latent infection within the first weeks or months of life.153 Establishment of latency with subsequent viral reactivation resulting in clinical disease and viral shedding during periods of stress is a characteristic of herpesviruses in general, including EHV-1 and EHV-4,121 and makes eliminating these pathogens from the equine population a difficult if not impossible endeavor. Latency and reactivation are critical features of the epidemiology of EHV-1 and EHV-4 infection, allowing the virus to persist for lengthy periods, and in all likelihood as a lifelong infection, in the majority of previously infected horses before reactivating. Reactivation and subsequent nasal virus shedding can result in infection of new susceptible horses and possibly in disease in the latently infected horse itself. Although the true prevalence of latent equine herpesvirus infection is unknown, as more sophisticated methods of detecting latent infection with EHV-1 and EHV-4 are developed,154 the detectable prevalence of infection continues to rise with the implication that EHV-free horses are a very rare occurrence.
■ Pathogenesis The pathogenesis of EHV-1 infection has been comprehensively reviewed.121,155,156 Primary EHV-1 or EHV-4 infection occurs at the respiratory epithelium, resulting in erosion of the upper respiratory mucosal surface and viral shedding for 4 to 7 days post infection; shedding can occur for longer at lower levels and possibly beyond 14 days. Cell-to-cell spread results in the presence of virus in respiratory tract lymph nodes within 24 to 48 hours post infection. In contrast to EHV-4 infection, which is mostly limited to the respiratory tract, EHV-1 establishes a lymphocyte-associated viremia, which is responsible for the delivery of virus to the pregnant uterus and other tissues. Spread of virus from leukocytes to endothelial cells at these secondary sites results in vasculitis, hemorrhage, thrombosis, and ischemic tissue necrosis. Viremia can persist for up to 21 days and is a prerequisite for abortion, chorioretinopathy, and EHM. Moreover, there appears to be a positive correlation between the development of EHM and the magnitude and duration of viremia.157 Despite this, only a small percentage of horses with viremia develops EHM, indicating that the clinical outcome of infection in the individual animal is influenced by other factors (e.g., age, sex, breed, virus strain).146 In pregnant mares, EHV-1 infection can result in abortion subsequent to vasculitis of the small arteriolar branches of the glandular layer of the endometrium with subsequent thrombosis, microcoty- ledonary infarction, perivascular cuffing, and transplacental spread of virus.121 Abortion typically occurs in the last 4 months of pregnancy but has been reported to occur as early as the second trimester. There are usually no signs of impending abortion, and the foal is typically expelled in the amniotic membrane, having just died from asphyxia due to placental separation. Some fetuses survive initially but then die from respiratory failure due to infection of the lungs. EHV-1 myelo- encephalopathy results from widespread vascular injury after damage to the vascular endothelium of the blood-brain barrier and thrombo-ischemic necrosis in the spinal cord and the brain. It is possible that the immunopathologic response to EHV-1 infection of the vascular endothelium of the central nervous system may play a part in the etiology of the thrombo-ischemic necrosis. In this regard, a recent study has found an increased concentration of D-dimers in the bloodstream during EHV-1 viremia, suggesting that viremia can result in activating the coagulation cascade and the fibrinolytic system.158 EHV-1 chorioretinopathy is also caused by a vascular endotheliopathy with subsequent ischemic injury to the chorioretina, and it is characterized clinically by focal or multifocal “shotgun” lesions.159 While the incidence of ocular lesions has been reported to be more than 50% following EHV-1 infection, most ocular infections remain subclinical.159 Moreover, these lesions are typically not detectable in acute disease and take weeks following infection to become visible. Latency is established in the lymphoreticular 121147160
system and in the trigeminal ganglion. ’ ’ Reactivation of latent virus frequently results in reinfection of the upper respiratory tract and shedding of virus in nasal secretions.
Immunity to EHV-1 and EHV-4 has been extensively reviewed.161 The protective immune response following infection is short lived (3 to 6 months), although this may increase with age and repeated exposure. High titers of virus-neutralizing (VN) antibody before infection can reduce the amount and duration of virus shed from the nasopharynx162 but fail to prevent infection or the development of EHM or abortion.163 The reason for this failure is likely to be related to the rapid intracellular translocation of the virus,164,165 thereby escaping the effects of VN antibody. However, intracellular virus is susceptible to cytotoxic T lymphocytes (CTLs), which can lyse virus-infected cells. This CTL activity is MHC class I restricted and mediated by CD8+ lymphocytes.166 Ponies with high frequencies of CD8+ CTLs show a reduction in the clinical and virologic outcome of infection167 and are protected against abortion.168 Control of EHV-1 infection requires an immune response that not only limits primary infection but also controls reactivation of latent infections.163 Although circulating and mucosal antibodies each play some role in protection,162,169 the most important component of a protective immune response to EHV-1 is the CTL response.168,170,171 Another important aspect of EHV-1 pathogenesis and immunity is the capacity of the virus to evade the immune response.172 Strategies of immunoevasion used by EHV-1 include modulation of cytokine responses with subsequent effects on T- and B-cell responses, modification of the CTL responses, interference with antigen presentation by downregulation of MHC class I expression, inhibition of antibody-dependent cytotoxicity, alteration of NK-cell lysis, and loss of efficient chemoattraction of professional antigen-presenting cells.173-176
■ Clinical Presentation Clinical diseases associated with EHV-1 and, to a lesser extent, EHV-4 infection include respiratory disease, abortion, EHM, chorioretinopathy, and neonatal foal death. Infection with both EHV-1 and EHV-4 causes respiratory signs, but clinical signs are likely more severe for EHV-1, as is the likelihood of viremia. Respiratory disease is often more severe in young horses than in older, previously exposed horses. Clinical signs of EHV-1 or EHV-4 respiratory tract infection include pyrexia, which may be biphasic, with a first peak at 24 to 48 hours associated with upper respiratory tract infection and a second peak occurring at 4 to 8 days if viremia develops. Lethargy and anorexia are often mild, and nasal discharge, serous at first, becomes mucopurulent by days 5 to 7. Conjunctivitis, lymphoadenopathy of the submandibular lymph nodes, and edema or vasculitis of the distal limbs are variable findings. Respiratory disease, particularly affecting a large number of animals, is uncommon in horses older than 2 years of age presumably due to recurrent exposure to the virus and development of sufficient immunity to suppress clinical signs of airway disease.
Neonatal foals infected with EHV-1 in utero may be normal at birth but quickly become weak, lethargic, and develop profound respiratory distress. Prognosis is poor for survival. The abortion and EHM forms of EHV-1 are discussed elsewhere under the appropriate system.
The clinical signs resulting from infection with EHV-2 and EHV-5 remain poorly described and established, although these viruses may be a cause of acute respiratory tract disease with clinical signs of nasal discharge, reactive submandibular lymph nodes, and coughing.123,1" !here is mounting support of a potential association of EHV-5 infection and EMPF. However, the exact nature of the relationship between the virus and the disease has not yet been fully elucidated.127,128,178-180
■ Prevention Due to the ability of equine herpesviruses to establish latency within their host, the elimination of these pathogens from the equine population is not a feasible approach to infection control. Therefore strategies that prevent dissemination of the virus from horses undergoing active virus shedding must be used. Vaccines are an attractive tool for this, but currently there are no products that can provide comprehensive protection against equine herpesvirus infection and its potential sequelae. Inactivated EHV-1 vaccines are capable of eliciting circulating VN antibody responses, which can limit respiratory disease and nasal viral shedding, and the use of some of these vaccines has been associated with a reduction in the incidence of abortion storms.161-163,181 Vaccines containing higher antigen loads, and typically those licensed for control of abortion, can generate high-titered VN antibody responses and even EHV-1-specific interferon-gamma responses.181 Such vaccines are likely to outperform other inactivated products with lower antigen loads. Using an optimized vaccination regime (vaccination was performed on days 0, 27, and 97) and virus challenge at the peak of the resulting immune response, a recent study was able to demonstrate a significant reduction in days of viremia in addition to a reduction of clinical signs and nasal virus shedding after administration of a commercially available high-antigen load killed vaccine.182 A commercially available modified live EHV-1 vaccine (MLV) generates limited EHV-specific cellular immunity183 and has been demonstrated to reduce respiratory disease and viral shedding but no protection against viremia.184 The use of current commercial vaccines of proven efficacy is recommended at 6-month intervals for control of respiratory disease and shedding of EHV-1 and EHV-4. Generally, vaccines effective against EHV-1 will cross-protect against EHV-4. There is no evidence that any commercial or experimental vaccine can prevent the occurrence of EHM.
No attempts are made to control EHV-2 or EHV-5 infection with vaccination. However, the proposal that concurrent EHV-2 infection may play a role in the etiology of R. equi pneumonia has led some clinicians to attempt vaccination of foals with killed EHV-2 vaccines.185
Biosecurity measures following detection of EHV-1 and EHV-4 infection should include the use of strict hygiene and barrier precautions as well as isolation of affected and in-contact horses for periods of up to 28 days. The virus is thought to spread primarily through contact, with limited airborne droplet spread compared to equine influenza virus. Extensive codes of practice have been described for control of the spread of EHV-1 in facilities both in publications121 and in online resources (http://www.hblb.org.uk; http://www.aaep.org).
Equine Arteritis Virus
Equine viral arteritis (EVA), named after the characteristic inflammatory changes found in the walls of small arteries, is a contagious disease of horses caused by equine arteritis virus (EAV).186 While EVA is a disease with worldwide distribution, the seroprevalence varies substantially between countries and equine populations. Despite the fact that the vast majority of EAV infections are subclinical, occasional outbreaks occur that result in respiratory disease in adult horses, abortion, and fatal pneumonia in foals. These disease outbreaks have significant costs in terms of animal suffering and economic loss. Recent examples of larger outbreaks with substantial impacts on animal health and economic losses include a multistate EAV outbreak in 2006-2007 in the United States as well as a 2007 outbreak in the Normandy region of France.187-189
■ Etiology EAV is an enveloped positive-strand RNA virus and is a member of the family Arteriviridae, order Nidovirales.190 The Arteriviridae comprise the following genera: Equartevirus (equine arteritis virus [EAV]), Rodartevirus (porcine reproductive and respiratory syndrome virus [PRRSV], lactate dehydrogenaseelevating virus [LDV] of mice), Simartevirus (simian hemorrhagic fever virus [SHFV] and others), and Dipartevirus (wobbly possum disease virus [WPDV]).191 The polycistronic genome of EAV is approximately 12.7 kb and contains at least 10 functional ORFs.192-194 ORFs 1a and 1b encode two replicase polyproteins that are posttranslationally processed by three proteinases to produce at least 13 nonstructural proteins. The remaining eight ORFs encode eight structural proteins, two (M and GP5) of which serve as the major viral envelope proteins. The GP5 envelope glycoprotein carries the major neutralization determinants of the virus, and four neutralization sites (A to D) have been identified.195 Based on phylogenetic analyses of the EAV-ORF5, the viruses are divided into two distinct clades: North American and European clades that share approximately 85% nucleotide identity.196,197 Despite the fact that only one serotype has been identified, there are extensive variations in virulence of different isolates of the virus. While the molecular determinants of EAV virulence have not been fully elucidated, recent data suggest that amino acid substitutions in the replicase and/or the structural proteins (GP2, GP4, GP5, and M) result in attenuation of a virulent strain of EAV.198 The virus is readily inactivated by lipid solvents and disinfectants, although it can remain infectious for 75 days at 4° C, for years in tissue samples stored at -20° C, and for decades in semen, embryos, and tissue samples stored at -70° C.199
■ Epidemiology EAV is spread by the respiratory and venereal routes, and the persistently infected carrier stallion is central to the persistence of the virus in the horse popula- tion.186 As a consequence, disease outbreaks frequently follow the importation of carrier stallions or their semen. With the increasing internationalization of the horse industry, this has become an important factor in the spread of the virus throughout the world in recent years and highlights the fact that identification of the carrier stallion is a key component of EVA control
187200
programs.18',200 Virus transmission may also occur through direct or indirect exposure of seronegative animals to aborted fetuses and fetal membranes, by carrier stallions to susceptible stallions by masturbation, and from EAV-infected donor mares to naive recipient mares via embryo transfer.201,202
Following infection, a variable percentage (10% to 70%) of stallions infected become carriers and shed the virus for a variable period, ranging from several months to years, in 200203
their semen.200,203 The virus persists primarily in the ampulla of the vas deferens, and the carrier state has been shown to be testosterone dependent, as castration resolves infection.200 Some carrier stallions spontaneously eliminate the virus from their reproductive tract. During replication of the virus in the carrier stallion, genetically and phenotypically distinct variants of the virus can emerge, and these variants subsequently can be found in the semen of these stallions.188,204,205 When one of these variants is transmitted to a mare through breeding, or occasionally through contaminated fomites, a new outbreak can occur.186 Transmission by the venereal route is associated with infection rates in mares as high as 85% to 100%, and disease outbreaks often result from breeding a seronegative mare with EAV-containing semen.190 Subsequent spread of the virus to other horses on the premises is by respiratory droplets and the airway route.190 During these outbreaks the EAV strain remains stable with minimal genetic change.186 Seroprevalence varies among breeds and between countries and geographic regions, with a reported seroprevalence of up to 85% in U.S. Standardbreds but less than 6% in Thoroughbreds in the United States.206 While it has been hypothesized that differences in seroprevalence are due to genetic susceptibility to EVA, it is likely that the difference is a consequence of the number of actively shedding carrier stallions in different breeds. The virulence of strains in specific breeds can vary, however, and the strains shed by Standardbreds are typically highly attenuated, causing minimal disease.186 Interestingly, recent research suggests that a correlation exists between the host's genotype (i.e., CD3+ T-cell susceptibility to EAV infection) and the clinical outcome of infection, including the potential to become a carrier of the virus.207-209
The immune response to infection is strong, leading to protective immunity with a duration of several years.190 Foals born to immunized mares are protected by colostral antibodies for 2 to 6 months.
■ Pathogenesis Virus transmission occurs via both the venereal and the respiratory routes. After respiratory infection the virus invades the respiratory epithelium and infects bronchial and alveolar macrophages, reaching bronchial and other regional lymph nodes by 48 hours post infection.190 Nasal virus shedding is typically detected for 7 to 14 days after infection but can last for up to 21 days.210 The establishment of viremia leads to dissemination of the virus throughout the body and further replication of the virus in the adrenals, thyroid, liver, and testes. Following infection, virus can be detected in the buffy coat for 2 to 21 days and in the plasma and serum for 7 to 9 days. The virus infects and replicates in endothelial cells, causing direct injury to the vascular endothelium and adjacent muscularis media, and subsequently gains access to the media of affected vessels. Vasculitis is characterized by fibrinoid necrosis of small muscular arteries, leukocyte infiltration, and perivascular hemorrhage and edema. With the exception of the carrier stallion, virus is eliminated by 28 days post infection and coincides with the appearance of virus-neutralizing antibodies.211 Reports exist where virus has been recovered for up to 180 days post infection from the accessory sex glands of prepubertal colts 190,212,213
■ Clinical Presentation The great majority of infected horses are asymptomatic. When disease does occur, the clinical signs of EAV infection can vary from fever and leukopenia to severe disease and death. In general, the severity of EVA tends to be greater in very young and old horses. The incubation period is 2 to 14 days (6 to 8 days following venereal exposure). Horses may show some of the following signs. Typically the affected animals are pyrexic, with temperatures reaching 40.5° C (105° F) for 1 to 5 days. Anorexia, lethargy, serous nasal and ocular discharge, and coughing may occur. Conjunctivitis (“pink eye”) with ocular discharge and periorbital and supraorbital edema; dependent edema of the limbs, prepuce, scrotum, and mammary glands; urticaria (localized to the sides of the neck and face or generalized); photophobia; petechial hemorrhage in the nasal, conjunctival, and oral mucous membranes; and submandibular lymphadenopathy have also been reported.190
In stallions, acute infection can cause short-term subfertility characterized by reduced libido, decreased sperm motility, sperm concentration, and normal sperm morphology. These changes in semen quality are transient but can persist for up to 16 weeks.187 In contrast, persistent infection does not appear to have an effect on sperm quality in carrier stallions.187
Abortions in pregnant mares occur in the acute or early convalescent phases of infection and are often not preceded by premonitory signs. It is thought that abortion occurs as a consequence of placental dysfunction secondary to vasculitis affecting myometrial blood vessels.214 Abortion can occur from 2 to 10 months of gestation, with abortion rates ranging from 10% to 70%. Fetuses are typically partially autolyzed at the time abortion occurs. Commingling of mares recently bred by an infected stallion with pregnant mares is a common scenario leading to abortion.
Neonatal foals infected with EAV show severe respiratory signs and high mortality. In foals that survive more than 24 hours, fever and leukopenia or thrombocytopenia are present. Interstitial pneumonia, lymphocytic arteritis or periarteritis, renal tubular necrosis, and fibrinoid necrosis of the tunic media are seen on pathologic examination.186
■ Prevention EVA control programs are aimed at eliminating the risk of abortion, neonatal foal death, and the establishment of the carrier status in stallions.199 Control measures include identification of carrier stallions, isolation and testing of horses prior to introduction into the resident horse population, segregation of pregnant mares and newborn foals, vaccination, and the establishment of biosecurity measures. Prevention of the development of carrier status in future stallions can be accomplished through vaccination of all colts with breeding potential in the first year of life and annual revaccination thereafter.190 Identification of the carrier stallion can be accomplished by virologic and serologic testing (taking into account previous vaccination history) and should be carried out prior to introducing a stallion to a new premises.199 Serologic testing should be performed in all stallions prior to the breeding season, and if seropositivity is detected in a stallion with no known vaccination history, virologic testing of the semen should be conducted. If the carrier status has been identified, these stallions should be managed separately under strict hygiene protocols to prevent spread of the virus to naive in-contact animals.187,200 Mares being bred to carrier stallions should be vaccinated against EAV. Fresh and frozen semen from carrier stallions is also infective, and negative mares receiving this semen should be vaccinated. Extensive codes of practice have been produced by both the U.S. Department of Agriculture (http://www.aphis.usda.gov) and the Horserace Betting Levy Board (http://www.hblb.org.uk) to assist in control of EVA. Moreover, guidelines for vaccination of horses against EAV have been established by the American Association of Equine Practitioners (AAEP; http:// www.aaep.org).
Both a modified live and a killed vaccine against EAV have been developed.186,190 In the United States, the MLV vaccine has been available since the mid-1980s. The vaccine was derived from an experimental vaccine that was created by serial passage of the virulent Bucyrus strain in primary horse kidney and primary RK cells. The MLV vaccine provides complete or partial protection against the clinical signs of EVA, but viral replication still occurs after challenge. Despite the fact that only a single serotype of EAV has been described, recent in vitro studies demonstrated that antibodies raised subsequent to vaccination with the MLV vaccine do not neutralize all strains with equal efficiency.215,216 The duration of vaccinal protection appears to be as long as 2 years, although annual revaccination of breeding stallions is recommended before the start of the breeding season. The MLV vaccine has been used successfully to limit outbreaks of infection in racecourses after initial diagnosis.190 The manufacturer does not recommend the use of this vaccine in foals younger than 6 weeks of age and in pregnant mares. Vaccination of pregnant mares during the last 2 months of gestation has been associated with the risk of abortion.217
Equine Rhinitis Virus
■ Etiology Equine rhinitis viruses (formerly equine rhinoviruses) are a known cause of clinical and subclinical respiratory infections in horses worldwide. The viruses are classified as members of the family Picornaviridae, which includes small (“pico”), nonenveloped, single-stranded RNA viruses. Among the equine rhinitis viruses, at least two serotypes have been isolated from horses: equine rhinitis A virus (ERAV) (formerly equine rhinovirus 1 [ERV-1]), and equine rhinitis B virus (ERBV) (formerly equine rhinovirus 2 [ERV-2]).218,219
Considerable uncertainty has surrounded the classification of the equine rhinitis viruses. Because the illness caused by these viruses resembles the common cold brought about by rhinoviruses in humans, they initially were included in the genus Rhinovirus. However, subsequent studies demonstrated that ERAV differed from other members of the Rhinovirus genus as well as from ERBV.219-222 For example, while rhinoviruses generally demonstrate a narrow host-cell range in vitro and in vivo, ERAV is able to infect and replicate in a broad range of cell types and host species. For instance, intranasal inoculation of a human volunteer with ERAV resulted in pharyngitis, lymphadenitis, fever, and viremia.223 Furthermore, the presence of systemic signs and viremia during ERAV infection, the occurrence of persistent infections, and the physical properties of the virus are all more reminiscent of foot-and-mouth disease virus (FMDV). Based on these characteristics, along with similarities in protein sequences, ERAV was placed alongside FMDV in the Aphtovirus genus of the picornavirus family. Subsequently, ERBV has been reclassified as the sole member of the new genus Erbovirus. To date there are three known serotypes of ERBV: ERBV1, ERBV2, and ERBV3 (acid-stable equine picornavirus).
■ Epidemiology First isolated from horses in the United Kingdom,224 ERAV infection occurs worldwide and is a recognized cause of acute respiratory disease.218,225-227 The incidence of neutralizing antibody to ERAV varies according to the age of the horse, and most horses appear to become infected during their second year of life. In a study by Studdert and Gleeson, only 16% of horses between the age of 6 and 12 months were seropositive.221 However, studies conducted in older horses found that a high percentage (50% to 90%) of horses 5 years of age have antibodies to equine rhinitis virus.225,228-230 As with many other respiratory pathogens, young horses entering a large population in frequent contact with other horses are most susceptible to infection.225 In addition, there appears to be a seasonal distribution, with most of the ERAV
231232
infections occurring in late winter and spring.231,232 Stress (e.g., transportation and training) and concurrent disease resulting 233
in immunosuppression may predispose to infection.233 The virus is fairly resistant to inactivation and is thought to survive in the environment for a considerable time. As for human rhinoviruses, virus spread occurs most likely through direct and indirect contact (fomites).234
The clinical significance of ERBV1, ERBV2, and ERBV3 is not well defined. Several recent surveillance studies indicate that the prevalence of ERBV1- and ERBV2-neutralizing antibodies is between 24% and 80% of horses.123,229,230,235 However, the consequences of infection with ERBV1 and ERBV2 are unclear, as these viruses have been isolated from horses both with and without signs of acute respiratory illness.229,236,237
■ Clinical Presentation Although ERAV is generally considered a respiratory pathogen, the clinical signs associated with infection may be quite variable. Following experimental infection with ERAV, horses may develop pyrexia, anorexia, nasal discharge, pharyngitis, and lymphadenitis involving at least the submandibular and pharyngeal lymph nodes.238 On some occasions, laryngitis or mild bronchitis can develop.228 Based on serologic studies, subclinical ERAV infection may also occur and may be important in the dissemination of the disease.221,231,234,238,239
Horses infected with ERAV may also develop viremia. On average, viremia lasts for 4 to 5 days, and the disappearance of virus from the blood coincides with the appearance of neutralizing antibodies.238 Although the relationship between antibody levels and protection against disease has not been established, immunity is thought to be solid, and antibodies
234
persist for a long time.234
Infection with ERAV may be accompanied by long-term urinary and fecal shedding as well as virus persistence.238,240 The recognition of the potential for ERAV persistence has important implications for disease diagnosis. Highly sensitive diagnostic methods (e.g., immunofluorescent antibody [IFA] or RT-PCR) may be able to detect persistent ERAV in horses suffering from respiratory disease, which may not be associated with the observed illness. This notion is underlined by the recent finding that the ERAV genome was detected by RT-PCR in nasopharyngeal swab samples obtained from two clinically healthy horses 1 year after acute ERAV infection.219
■ Control Similar to other respiratory viruses, infection control should be based on adequate husbandry and biosecurity measures. Recently, a conditionally licensed ERAV vaccine has become available in the United States. Based on data provided by the manufacturer, the initial vaccination series should include three doses administered with a 3- to 4-week interval between each dose. Booster vaccinations should be administered at yearly intervals or prior to an anticipated virus exposure.
Equine Adenovirus
■ Etiology Adenoviruses (family Adenoviridae) comprise a group of nonenveloped, icosahedral DNA viruses. Although most vertebrate species harbor an adenovirus, the viruses are highly species specific. In humans, adenoviruses are extremely prevalent and represent a significant cause of acute respiratory, ocular, and GI disease. Infections of immunocompetent individuals usually stay localized and often take a mild or subclinical course.241 Similarly, equine adenoviruses have been isolated from clinically healthy adult horses242 and foals.243,244 Equine adenoviruses consist of two species, equine adenovirus 1 (EAdV1) and equine adenovirus 2 (EAdV2), which appear to have evolved from separate lineages within the adenovi- ruses.245 EAdV1 has been implicated as the cause of acute respiratory tract disease246,247 and conjunctivitis, as well as a progressive, fatal pneumonia in Arabian foals with SCID.248-250 EAdV2 has been incriminated as a cause of intestinal infec- tion251,252 and diarrhea in immunocompetent foals.253,254
■ Epidemiology First isolated in 1969 from a foal with respiratory disease,255 equine adenoviruses have a worldwide distribution.256 While serologic evidence from Australia, the United Kingdom, the United States, the Netherlands, and Nigeria indicates that adenovirus infection occurs at a high frequency in the general horse population,257-260 the role of equine adenovirus in respiratory disease in adult horses remains unclear. In this regard it is interesting that a number of studies found no apparent association between clinically apparent respiratory disease in young adult horses and infection with adenovirus, as diagnosed by subsequent seroconverison.237,239,261
In contrast, fatal and nonfatal adenovirus pneumonia has been well documented in foals, particularly of the Arabian breed. Fatal respiratory disease is predominantly seen in foals with SCID. Other foals may develop respiratory disease but usually do not succumb to disease.
In general, adenovirus transmission is thought to occur via direct contact with secretions from infected animals or indirectly via fomites or infected premises. In adult horses, the virus can persist in the upper respiratory tract, and these animals can act as a reservoir host for the virus. Adenoviruses are highly stable and have been found to remain infectious at 4° C for up to 1 year.
■ Clinical Presentation The clinical signs recorded after experimental intranasal infection of a Thoroughbred yearling were confined to a serous nasal discharge that lasted from 4 to 12 days post inoculation. Peak levels of serum antibodies were found at 13 days after inoculation and dropped by approximately 2 months after experimental infection.262
In foals, clinical signs generally occur between 10 and 35 days of age.263 After experimental inoculation at 24 hours of age, clinical signs appeared 3 to 5 days after infection and consisted of intermittent pyrexia, nasal and ocular discharge, tachypnea, and cough. All foals recovered by day 10 post inoculation. The majority of foals developed a mild and transient lymphopenia that was followed by lymphocytosis. Typically, about 25% of infected foals also develop diarrhea.263 Gross pulmonary lesions found in animals euthanized at 14 to 21 days post inoculation included atelectasis, involving up to 25% of the pulmonary parenchyma, and suppurative bronchopneumonia. Histologically, swelling and hyperplasia of the airway epithelium, as well as intranuclear inclusion bodies in the respiratory epithelial cells, were a consistent finding up to 11 days post infection.264
Foals with SCID show a progressive decline in health despite intensive therapy. In necropsy, conjunctivitis, rhinitis, tracheitis, and bronchopneumonia are present. Intranuclear inclusion bodies are readily demonstrated in the respiratory epithelial cells. Areas of interstitial pneumonia surround affected bronchi. Acute pancreatitis characterized by inclusion bodies in acinus and ductal cells, as well as adenoviral sialoadenitis involving labial and parotid salivary gland can often be demonstrated.252,265
■ Control No vaccine is available to control adenovirus infection in horses. However, except in immunocompromised animals, the virus appears to be of little clinical importance.
Hendra Virus
Hendra virus is described in detail later in this chapter.