Overview of Immunologic Disorders in Horses
Lais R.R. Costa
The immune system is composed of primary or central organs (bone marrow and thymus) and secondary or peripheral organs (spleen, lymph nodes, and mucosa-associated lymphoid tissues).
Immune cells of myeloid and lymphoid lineages are generated (produced and developed) in central organs, subsequently populating the peripheral organs where they carry out their functions of immunosurveillance and mounting immune responses to defend the body from microbial invasion. Many immune cells are not static; instead, they are constantly circulating, migrating into various tissues, and capable of performing their functions of immunnosurveillance, inflammation, and tissue repair in organs outside the immune system. Functionally, immune cells traditionally have been classified into two branches: innate and adaptive immune cells. Cells of the innate branch of the immune system mostly comprise cells of the myeloid lineages—including granulocytes (neutrophils, eosinophils, basophils, mast cells), monocytes and macrophages, dendritic cells (DCs)—and subsets of cells of the lymphoid lineage—including natural killer (NK) cells, plasmacytoid DCs, B1 cells, and gamma delta T cells. Cells of the adaptive branch of the immune system primarily comprise cells of the lymphoid lineages expressing antigen-specific receptors, including B lymphocytes, T lymphocytes (alpha beta T cells of either CD4 and CD8 subpopulations), and regulatory T cells. The various components of the immune system work in unison to bring about broad defense throughout the body. Detailed review of the cellular and molecular components of the immune system and the mechanisms involved in the immune response of horses can be found elsewhere.1-3Normal Immune Response in Horses
The immune response results from an intricate network of innate mechanisms of resistance and complex adaptive immune defense.
The innate immune mechanisms are the first line of defense and do not require previous exposure to the invader. Cell-associated molecules and soluble factors of the innate system are capable of recognizing a number of common microbial structures or patterns and respond quickly to an invasion by promoting inflammatory reaction and chemotactic stimuli for leukocyte trafficking, thus serving as the first immune barrier to invading organisms. Cells and soluble factors of the innate system also provide co-stimulatory signals and antigen presentation to cells of the adaptive lymphoid lineage. Once mounted, the adaptive host defense confers antigen-specific anamnestic response following the initial encounter with an antigen or invader. The adaptive immune response can recognize nearly an infinite array of antigenic determinants or epitopes, distinguishing self from those that are nonself. As well, the adaptive immunity has memory that leads to increased effectiveness upon antigenic reexposure. Thus the innate and adaptive mechanisms work together to identify and eliminate offending molecules and organisms that invade the body.Innate recognition of invaders involves detection of alarm signals generated exogenously (invader patterns) or endogenously (patterns of damaged cells or tissue). Several cells of the innate immune system, including cells of the macrophage lineage, DCs, mast cells, neutrophils, NK cells, and gamma delta T cells, as well as B cells can recognize exogenous and endogenous alarm signals. Sentinel cells scattered throughout the body, especially at sites likely to be invaded (skin and mucosal surfaces), serve the primary function of invasion recognition. Exogenous invasion signals (called pathogen-associated molecular patterns [PAMPs]) and endogenous markers of danger (called damage-associated molecular patterns [DAMPs]) are recognized by pattern recognition receptors (PRRs) expressed on macrophages, DCs, and mast cells. Typically PAMPs are conserved structural molecules that are abundant and commonly found in different organisms.
Examples of extracellular and intracellular PAMPs are depicted in Fig. 53.1, A. Alarmins, another name for DAMPs, may be of intracellular origin when released upon cell death or of extracellular origin when released upon damage of connective tissue (Fig. 53.1, B). PRRs are generally expressed on cell membranes (for recognition of extracellular pathogens) or within the cytosol and cytoplasmic vesicles (for recognition of intracellular pathogens), but some PRRs circulate in blood as soluble factors (Fig. 53.1, C). A number of families of PRRs have been identified, with Toll-like receptors (TLRs) being one of the most important families. The binding of a signature PAMP or DAMP to a particular PRR activates a signaling pathway that results in transcription and translation of specific genes characteristic of an appropriate response against the triggering pathogen. Some of the known PRRs and the corresponding PAMPs and DAMPs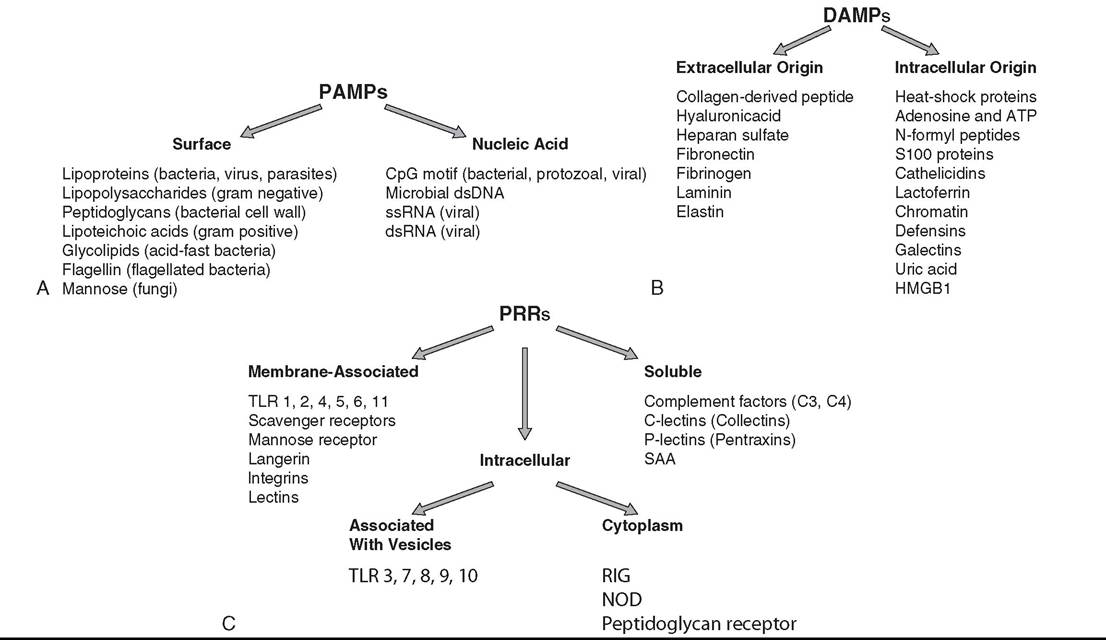
FIG. 53.1 A, Pathogen-associated molecular patterns (PAMPs) are exogenous invasion signals that comprise conserved and essential molecules abundantly and commonly found in many different organisms. CpG, Unmethylated cytosine-phosphate-guanine deoxynucleotide; dsDNA, double-stranded deoxyribonucleic acid; dsRNA, double-stranded ribonucleic acid; ssRNA, single-stranded ribonucleic acid. B, Danger-associated molecular patterns (DAMPs) are endogenous invasion signals that are released upon cell death or damage of connective tissue. ATP, Adenosine triphosphate; HMGB1, high mobility group box 1 protein. C, Pattern recognition receptors (PRRs) recognize pathogen-associated and damage-associated molecular patterns as signals of invasion and danger. NOD, Nucleotide-binding oligomerization domain-like receptor; RIG, retinoic acid-inducible gene 1 protein; SAA, serum amyloid A; TLR, Toll-like receptors.
they recognize and their signaling cascade are discussed in Chapter 32 and found elsewhere.2,4-7
Once alarm signals are recognized and cell signaling is activated, a cascade of events leads to development of an inflammatory reaction characterized by increased blood flow, increased vascular permeability, and leukocyte migration and activation.
Inflammatory response is reviewed in detail in the Endotoxemia and Sepsis section in Chapter 32. Soluble mediators of the innate immune mechanisms include acutephase proteins, proinflammatory cytokines, chemokines, vasoactive mediators, antimicrobial peptides, and protease systems (Table 53.1). Of note is the complement system activated upon specific immunologic recognition by antibodies (classical pathway), upon association of soluble factor C3b to microbial molecules (alternative pathway), or upon binding of soluble PRRs (lectins) to microbial carbohydrates. Complement activation leads to an amplifying cascade of sequential proteolytic cleavages, resulting in enhancement of phagocytosis (opsonization) and release of potent proinflammatory factors (C3a and C5a) that increase vascular permeability, exert chemoattraction of leucocytes, and amplify inflammation. The final product of the complement sequential proteolytic cleavage is the formation of lytic multiprotein complex, the terminal complement complex, or membrane attack complex.Neutrophils and macrophages take the center stage in innate immunity, as they are capable of destruction of offending organisms by phagocytosis, oxidative burst activity, and production of lytic enzymes. Neutrophils (polymorphonuclear granulocytes) are abundant and the predominant circulating leukocyte in horses. They are produced in the bone marrow at a very high rate; circulate for a short time (average of 12 hours); exit the bloodstream into capillaries of the liver, spleen, lungs, and bone marrow; and eventually migrate into tissues where they undergo apoptosis. Apoptotic neutrophils are removed by macrophages. Upon inflammatory stimulation, circulating neutrophils are mobilized via diapedesis, and production of new neutrophils in the bone marrow is induced in order to fulfill the recruitment demands, as neutrophils are terminally differentiated and do not divide. Neutrophils have three major types of enzyme-rich granules in their cytosol.
Upon stimulation, granules containing complex mixtures of bactericidal molecules are exocytosed. Release of enzymes mediates not only microbial destruction but also tissue damage. Once activated, neutrophils are the most abundant first line of phagocytes at sites of acute inflammation or infection.Macrophages originate from monocytes and become specialized phagocytic cells resident in various tissues, collectively called the mononuclear phagocyte system (formerly known as the reticuloendothelial system). Macrophages are named according to the tissues they home: microglial cells in the brain, histiocytes in connective tissue, Kupffer cells in the liver, splenic macrophages in the spleen, alveolar macrophages in alveoli of the lungs, pulmonary intravascular macrophages in capillaries of the lungs, and osteoclasts in bones. The mononuclear phagocyte system plays a critical role in removal of bloodborne pathogens and particles and of parasitized and aged red blood cells (RBCs) from the bloodstream when the blood passes through capillary beds lined by macrophages in
■ TABLE 53.1
Examples of Soluble Mediators of the Innate Immune System and Their Main Functions
| Categories of Mediators | Examples of Soluble Mediators | Functions |
| Acute-phase proteins | C-reactive protein | Opsonization, complement activation, soluble PRRs |
| Serum amyloid A | Leukocyte recruitment and activation, soluble PRRs | |
| Cerulloplasmin | Iron sequester (limiting nutrients required by | |
| Ferritin | microorganisms) | |
| Fibrinogen | Hemostasis | |
| Complement C3b | Factors of complement, involved in activation of | |
| Complement C4b | leukocytes, soluble PRRs | |
| Protease systems | Complement | Mediate inflammation, opsonization and MAC formation |
| Kinins | Mediate inflammation | |
| Clotting and fibrinolysis proteins | Hemostasis | |
| Host defense peptides and | Equine beta defensin | Antimicrobial (bacteriostatic and bactericidal) |
| enzyme | Equine neutrophil antimicrobial peptide Equine cathelicidin Lysozyme | actions |
| Type I interferons (IFN-α, IFN-β) | Antiviral glycoproteins mainly produced by macrophages, DCs, and fibroblasts | |
| Vasoactive mediators | Biogenic amines (histamine, serotonin) | Vascular smooth muscle relaxation (vasodilation) |
| Peptides (kinins) | and endothelial cell contraction (leakage of | |
| Lipids (PGs, TX, PAF, LTRs) | plasma into tissues) | |
| Proinflammatory cytokines | CXCL8 (formerly known as IL-8) | Chemoattractants for cells of both innate and |
| and chemokines | CCL2 (formerly known as MCP-1) | adaptive branches of the immune system |
| IL-1 | Mediate a variety of cellular responses such as | |
| IL-6 | chemotaxis of leukocytes, activation of leukocytes | |
| TNF-α | (including T and B cells), upregulation of iNOS and COX-2, and induction of fever | |
| Prostanoids | PGs LTRs PCs | Mediate and modulate acute inflammation |
| Lectins | P-lectins (pentraxin) | Bind microbial glycoproteins and polysaccharides |
| S-lectins (galectins) | Mediate binding of leukocytes to extracellular matrix | |
| C-lectins (dectins) | Surface PRRs |
COX, Cyclooxygenase; DCs, dendritic cells; IFN, interferon; IL, interleukin; iNOS, inducible nitric oxide synthase; LTRs, leukotrienes; MAC, membrane attack complex; MCP1, monocyte chemoattractant protein-1; PAF, platelet-activating factor; PCs, prostacyclins; PGs, prostaglandins; PRRs, pattern recognition receptors; TNF, tumor necrosis factor; TX, thromboxane.
the liver, spleen, and lungs.
The mononuclear phagocyte system is also responsible for the removal of apoptotic cells in tissues. Macrophages also function as sentinel cells responsible for recognition of invaders, release of alarmins, and production of mediators that amplify inflammation and promote neutrophil recruitment. Macrophages serve as a backup to neutrophils, being slower responders with greater phagocytic and antimicrobial abilities, particularly effective against intracellular organisms. Macrophages have other critical immune functions, including antigen processing and presentation to T cells, secretion of cytokines that stimulate lymphocytes and shape the adaptive immunity, and modulation and suppression of the immune response. Hence subpopulations of macrophages exert different immunologic functions. For instance, after exposure to PAMPs and interferon gamma (IFN-γ), macrophages differentiate into M1 cells that are potent amplifiers of inflammation, secreting large quantities of proinflammatory cytokines and producing large amounts of lysosomal enzymes and mediators involved in oxidative burst. In contrast, differentiation into M2 cells occurs after exposure to interleukin (IL)-4, IL-10, and IL-13, leading to anti-inflammatory response characterized by modulation of inflammation and promotion of tissue repair.DCs comprise a heterogeneous population of immune cells, originating from either myeloid or lymphoid precursors and exerting three main immune functions: serving as sentinel cells for the detection and recognition of invaders; providing antigen presentation to T cells, thus initiating the adaptive immune response; and guiding the type of adaptive response by regulating the milieu of co-stimulatory cytokines. Sentinel DCs are prominent at skin and mucosal surfaces. With respect to their origin, DCs are classified as DC1-type cells, which include myeloid-DC and Langerhans cells, originated from monocytes; or DC2-type cells, the plasmacytoid DCs that originate from lymphoid precursors. Myeloid DCs can convert to macrophages, and vice versa, and are considered part of the mononuclear phagocyte system. Plasmacytoid DCs are specialized in secreting type I interferons (IFN-α and IFN-β) upon viral infections, which activate lymphocytes and NK cells. DCs, along with macrophages, are the principal antigen-processing cells providing stimulation of T cells. Antigen processing and presentation in the context of major histocompatibility complex (MHC) classes I and II are critical functions linking the cells of the innate immune system and those of the adaptive immune system.
NK cells, gamma delta T cells, and B1 cells are lymphoid cells considered to be in the interface between innate and adaptive immunity. NK cells recognize cells under stress, including viral-infected or tumor cells that do not express antigenic epitopes in the context of MHC class I. Many viral- infected and tumor cells actively promote downregulation of MHC class I expression, which allows them to evade detection by cytotoxic T cells, also known as CD8+ alpha beta T lymphocytes (see below). In the absence of MHC class I molecule expression, the NK cells become activated and secrete perforins and granzymes that destroy the target cells. Another mechanism used by NK cells to destroy viral-infected and tumor cells is called antibody-dependent cell-mediated cytotoxicity (ADCC), which involves binding of receptors for the gamma heavy chain of immunoglobulin G (IgG) (Fc-gamma receptor) on NK cells to IgG bound to viral-infected or tumor molecules expressed on target cells. Once activated, NK cells release cytotoxic granules, containing granzymes and pore-forming protein perforin, onto the surface of the target cell, causing programmed cell death of the target cell. Gamma delta T cells comprise a heterogeneous population of T cells that express gamma delta T-cell receptors (TCRs) and exert a wide range of immune functions, including response to heat shock proteins, cytokine production, activation independent of MHC, and participation in delayed hypersensitivity reaction. In horses, the role of gamma delta T cells has not been well described. A subpopulation of B cells known as B1 cells has been partially described in the horse, and they are abundant during fetal and neonatal periods; with age, these cells concentrate in the peritoneal cavity. Equine B1 cells have diverse immunoglobulin light chain repertoires but more frequently express a receptor that binds to phosphorylcholine commonly present on bacteria, helminths, apoptotic cell membranes, and oxidized lipids; B1 cells are known to become activated, differentiate, and secrete antibodies in a T cell-independent manner but their origin and function still need to be determined in the horse.8,9
The adaptive (antigen-specific, clonal) immune response involves two major populations of lymphocytes, B cells and T cells (Fig. 53.2, A). B lymphocytes populate germinal centers of the spleen and lymph nodes and comprise 15% to 30% of the peripheral blood lymphocytes. Alpha beta T lymphocytes populate the periarteriolar regions of the spleen and the paracortical regions of lymph nodes and comprise about 70% to 80% of peripheral blood lymphocytes of horses.
In the adaptive immune response, the highly specific antigen recognition is mediated by B-cell immunoglobulin receptors (BCRs) on B lymphocytes and by alpha beta T-cell receptors (TCRs) on T lymphocytes. The diversity of these receptors, both BCRs and TCRs, that allows recognition of an enormous number of antigenic epitopes is generated by several mechanisms, including DNA recombination of different gene segments V, D, and J encoding the antigen-binding region, addition of nucleotides during the recombination process (junctional diversity), and point hypermutations. Both BCRs and TCRs are heterodimers, which in turn provides yet another mechanism to increase antigen-recognition diversity.
The B lymphocytes produce and express antibodies or immunoglobulins (Igs) on the cell surface and are the precursors of plasma cells, which secrete Igs. B cells mediate humoral immunity, which classically provides defense against toxins, extracellular bacterial infections, parasites, and certain viral infections. There are several isotypes of Igs, classified according to their heavy chain constant region—IgA (alpha), IgD (delta), IgE (epsilon), IgG (gamma) and IgM (mu heavy chain)—and each isotype confers special functions.10 For instance, dimeric IgA (joined by J chain) produced in mucosa-associated lymphoid tissues is the most abundant Ig isotype in body secretions, whereas monomeric IgA is present in serum; the main IgA functions are antigen neutralization and opsonization. IgD is a surface receptor of naive mature B cells, and its function is not well determined. IgE is present in small amounts in serum, and it is mostly found bound to Fc epsilon receptors on the surface of mast cells and basophils; when two or more IgE molecules bind to the antigen or allergen, it promotes cell degranulation. IgM is the original surface receptor of developing B cells and is present in naive and some activated and memory B cells; when secreted by plasma cells, IgM circulates as pentamer, and it promotes antigen neutralization and complement activation. IgG is the main Ig isotype in plasma and serum; IgG's main functions are neutralization and opsonization, and some subclasses promote complement activation. The characteristics and immunologic functions of the equine Ig isotypes and subclasses are summarized in Table 53.2.11-15 Besides antibody production, B cells function
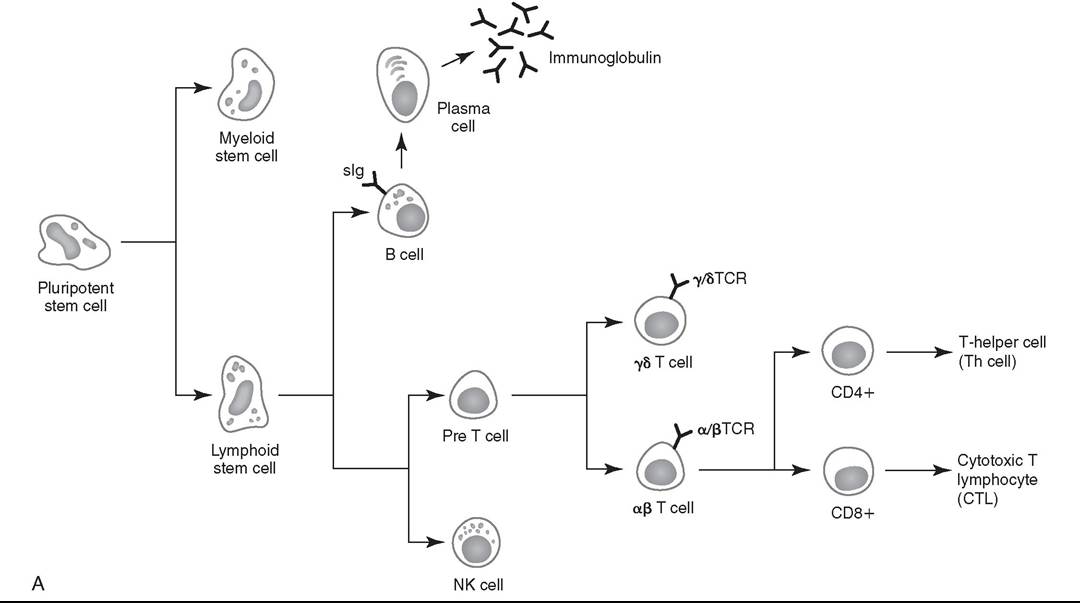
FIG. 53.2 A, Ontogeny of the lymphoid cell lineage. NK cell, Natural killer cell; sIg, surface immunoglobulin; TCR, T-cell receptor.
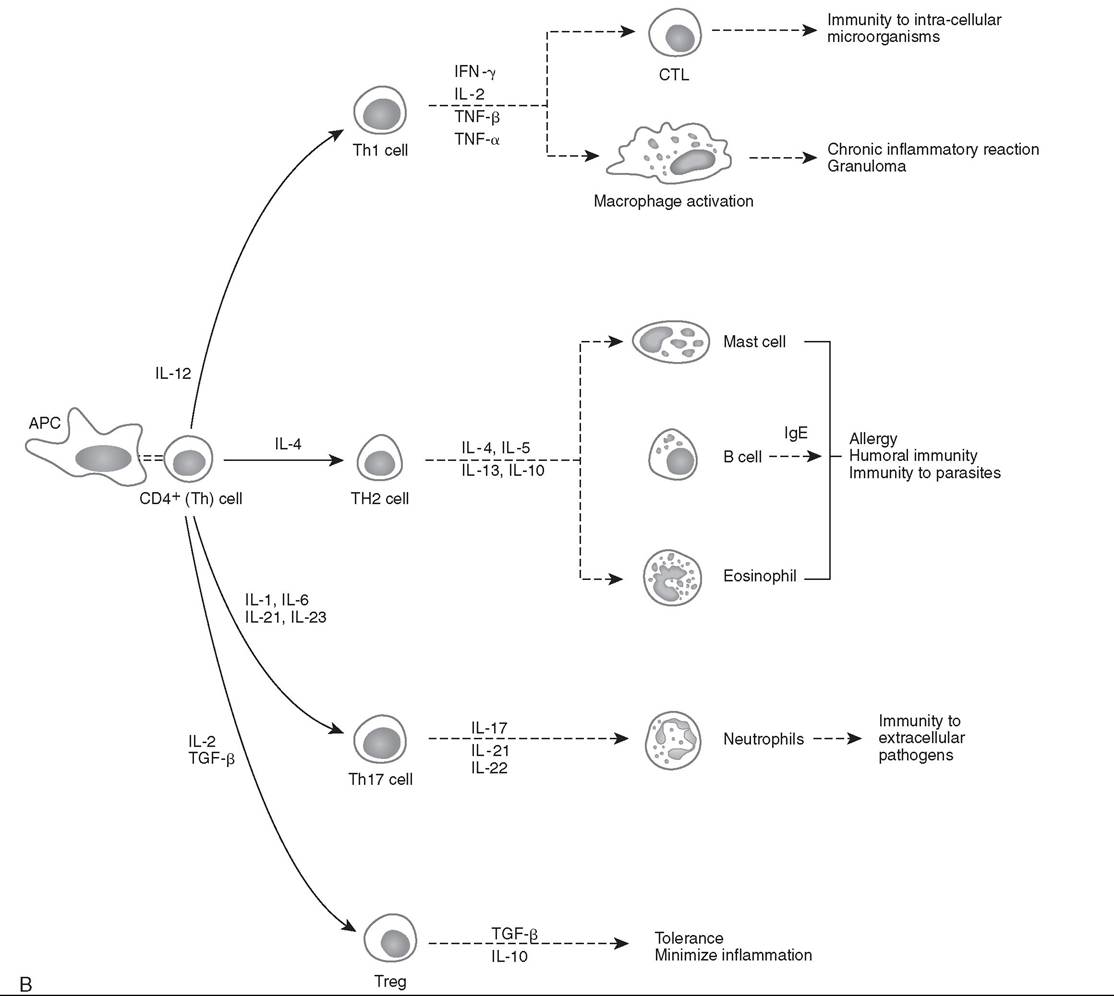
FIG. 53.2, cont'd B, Simplified functional differentiation of T-helper (Th) cells. CD4+ Th cells can be divided into subsets of Th cells based on their cytokine profile. Antigen-presenting cell (APC) interaction with Th cells via antigen presentation, co-stimulatory molecules, and cytokine milieu leads to the functional differentiation of subsets of Th cells. The cytokine milieu by APCs that leads to the functional differentiation of individual Th cells is listed. The known roles of the various functional subtypes of Th cells are also listed. CTL, Cytotoxic T cell; IFN, interferon; Ig, immunoglobulin; IL, interleukin; TGF, transforming growth factor; TNF, tumor necrosis factor; Treg, regulatory T cell;
as important antigen-presenting cells (APCs) by recognizing, internalizing, processing, and finally presenting antigens in the context of MHC class II to T cells. A less appreciated function of subsets of B cells is the production of cytokines. Subsets of B cells are now recognized as regulators of the immune responses during inflammation and autoimmunity.
T lymphocytes expressing alpha beta TCRs orchestrate the adaptive immune responses and provide cell-mediated immunity for protection against fungal, protozoal, intracellular bacterial, and viral infections. Unlike B lymphocytes, alpha beta T lymphocytes only recognize antigen fragments associated with the MHC molecule. The alpha beta T lymphocytes are categorized according to the expression of cluster of differentiation (CD) as CD4 or CD8. Functionally, CD4+ T lymphocytes comprise the T-helper (Th) cells, whereas CD8+ T lymphocytes comprise cytotoxic T cells (CTLs). The CD4+ T lymphocytes recognize antigenic epitopes in the context of MHC class II, which means that endocytosed antigens (e.g., extracellular pathogens) are processed and presented by APCs via the exogenous pathway. The CD8+ T lymphocytes recognize antigenic epitopes in the context of MHC class I, which means that the antigens are derived from intracellular proteins (e.g., intracellular pathogens) via the endogenous pathway. Activation of CD8+ T lymphocytes to become effector CTLs requires specialized APCs to be capable of cross-presentation, where antigen peptides that originated from exogenous foreign proteins are presented on MHC class I molecules.16 Specific subsets of DCs have adapted their endocytic and phagocytic pathways to
Characteristics and Immunologic Functions of Equine Immunoglobulin (Ig) Isotypes IgM1 IgG1 IgA1 and IgE and Subclasses IgGl to lgG7

Summary of data compiled from several authors (see references).
Concentrations of Ig isotypes and subclasses are approximated with the goal of highlighting the differences between the various isotypes.
Note that the Ig concentrations are given in mg∕mL. Most laboratories report Ig concentrations in mg∕dL. Conversion from mg/mL = 100 X mg∕dL. ADCC, Antibody-dependent cell-mediated cytotoxicity; MC, mast cell; NA, not applicable; NKs, natural killer cells; U, unknown.
acquire exogenous antigens from infectious agents and present them in the context of MHC class I, without being infected by the infectious agent.
Based on the profile of cytokine expression, CD4+ Th cells are classified into distinct functional subsets of Th cells that modulate the humoral and cellular immune responses. Fig. 53.2, B illustrates a simplified version of the functional differentiation of Th cells. APC interaction with Th cells via antigen presentation, co-stimulatory molecules, and cytokine milieu leads to the functional differentiation of particular subsets of Th cells. The secretion of IL-12 by APCs leads to the generation of the T-helper type 1 (Thl) cell subset, which produces primarily IFN-γ and functions in cell-mediated immunity, immunity to intracellular organisms, chronic inflammation, and delayed-type hypersensitivity (DTH) reactions. The cytokine IL-4 generates the T-helper type 2 (Th2) cell subset, which secretes IL-4, IL-5, IL-10, and IL-13 and interacts with B cells for the production of antibodies. The subset designated Th17 is induced by a cytokine milieu that includes IL-1, IL-21, IL-23, and IL-6; Th17 cells secrete proinflammatory cytokines IL-17, IL-21, and IL-22, which promote immunity to extracellular pathogens (bacteria and fungi).17,18 The immunomodulatory cytokine milieu based on IL-2 and transforming growth factor beta (TGF-β) promotes differentiation to the subset designated regulatory T cells (Tregs), which secrete the immunosuppressive cytokines TGF-β and IL-10. A number of other Th subsets have been described in human and animal models of disease, demonstrating the complexity of adaptive immune response.
Factors Associated With Adequacy of the Immune Response
Many factors influence the adequacy of the immune response of an individual animal to potential environmental organisms. Important factors to consider in the horse are age; genetic predisposition; exercise fitness; stress associated with exercise or transportation; state of health, including nutritional status; and the presence of concurrent diseases, including viral infections, tumors, and autoimmunity. Moreover, increased concentrations of corticosteroids can result in varying degrees of immunosuppression and may exacerbate preexisting infectious diseases or decrease resistance to environmental pathogens.19 The increase in endogenous corticosteroids may be associated with stress or disease; the most common condition in horses is caused by pituitary pars intermedia dysfunction (PPID).20 In addition, corticosteroid treatment is the most common iatrogenic cause of immunosuppression (see the Drug-Induced Immunosuppression section later in this chapter).21
Age has been shown to affect immune function in multiple species. Altered immune function has been identified at both ends of the age spectrum, in foals and older horses. Detailed overviews of the immune responses in young horses and older horses have been reviewed recently, and they are discussed in detail later in this chapter.22-29 At birth foals have no circulating Igs of their own, and they normally receive maternal transfer of colostral antibodies soon after birth. Colostrum is critical for neonatal immunologic protection. Although foals are immunocompetent at birth, their immune system is naive. Heathy neonatal foals appear to have neutrophilic phagocy- tocytic ability and killing capacity via oxidative burst activity comparable to that of mature horses.29-31 Differences in adaptive immune responses between foals and adult horses are not completely understood.32-35 Despite the apparently adequate preparedness of the immune system in young foals, their ability to develop a humoral immune response is believed to be affected by the maternal-derived antibodies. As the foals age, the maternal antibody levels decline and no longer provide comprehensive protection, but at the same time they may interfere with the foals' ability to mount an immune response. Aging is associated with changes in immunologic status, in particular a decline in the functionality of the immune system characterized by a chronic, low-grade proinflammatory state and a decreased antigen-specific immunity or clonal exhaustion.20,28 The aging process affecting the immune response is often referred to as immunosenescence.
Nutritional status is well recognized to exert a major impact on immune function, and nutritional deprivation produces impairment of the immune response.36 Collectively called nutritionally acquired immunodeficiency syndrome (NAIDS), these immunologic dysfunctions include protein-calorie malnutrition and deficiencies of single nutrients. Protein-calorie malnutrition and deficiencies of single nutrients that assist in nucleic acid metabolism lead to dysfunction of the adaptive immune response, especially the T-cell functions, and result in atrophy of lymphoid tissues. In these individuals, the cell- mediated immunity and humoral response to T cell-dependent antigens are subnormal. Deficiency of single nutrients can impair production of key proteins by lymphocytes as well as phagocytic cells. Inadequate nutritional status may result from
(1) increased and unmet nutritional requirement, including during growth, pregnancy, lactation, aging, and intense work;
(2) disease state associated with altered nutrient intake, such as conditions leading to hypophagia or anorexia; and (3) disease state associated with altered absorption of nutrients.37 Young, growing animals are most likely to develop NAIDS, followed by geriatric patients, contributing to the greater risk of infections in both age groups.
Exercise represents not only an increased nutritional requirement but also a physiologic stress that can significantly alter the immune response. However, defining the precise effects of exercise on the equine immune system and susceptibility to disease has been difficult because of the complexity of the immune system, host factors (e.g., age, level of fitness), and the variable nature of exercise. In general, it appears that exercise may have both positive and negative effects on the immune response.38-45 Suppressive effects, such as a decline in the CD4+/ CD8+ ratio of T cells, decreased lymphoproliferative responses, and suppression of the innate immune system, have been associated with strenuous high-intensity exercise, prolonged exhaustive exercise, or overtraining. In contrast, moderate exercise tends to have beneficial effects on the host defense mechanisms. Data in horses directly linking exercise-induced immunosuppression and increased susceptibility to infectious disease are limited. However, the potential immunosuppressive effects of exercise need to be recognized. Influenza infection in trained horses subjected to moderate exercise led to increased signs of clinical disease, but the duration of disease was unaffected.46 Unconditioned ponies vaccinated with a killed influenza vaccine and subjected to 5 days of strenuous exercise had an increased susceptibility to clinical influenza after challenge exposure compared with rested ponies.47 However, ponies with exercise-induced immunosuppression responded to the administration of an intranasal modified-live equine influenza vaccine and were protected from challenge.48
Immunologic Disorders Resulting From Compromised or Deficient Immune Responses
Increased susceptibility to infections, because of recurrence, chronicity, or severity of infectious illness beyond what is considered “normal,” raises the suspicion of inadequate immune response. Inadequate immune protection may result from immunodeficiency disorders or a state of immunosuppression.49-53 General clinical features associated with immunodeficiency disorders or immunosuppression include (1) repeated episodes of infections that respond poorly to standard therapy or return following discontinuation of antibiotic therapy; (2) infections associated with opportunistic or low pathogenicity organisms rarely observed in immunocompetent individuals; (3) systemic illness after administration of attenuated live vaccines; (4) failure to respond to vaccination; and (5) persistent marked abnormalities in leukocyte numbers. Pathogens most often associated with immunodeficiencies in horses include Pneumocystisjiravecii, Candida spp., Aspergillus spp., adenovirus, Cryptosporidium, and Rbodococcus equi in adult horses. The type of organism causing infection often suggests the type of defective immunity. When inadequate immune response is suspected, it is important to determine if there is a primary immunodeficiency disorder or a secondary state of immunosuppression.
Primary immunodeficiency disorders recognized in horses are characterized mainly by defects of components of the adaptive immune response and are outlined in Table 53.3. Yet it is possible that many forms of impaired or inadequate immune response in foals and mature horses are not clinically recognized or still poorly defined. Detailed information on the diagnosis and management of immunodeficiency disorders in horses are provided later in this chapter and found elsewhere.54
Inherited forms of complement dysfunction or factor deficiency have not been reported in the horse. However, depletion of complement component C3 associated with chronic infections, such as equine infectious anemia, has been reported. Moreover, the foal is born with low serum complement activity, and colostrum is not a significant source of complement. Therefore foals are transiently deficient in serum opsonic capacity and rely on their own production of complement after birth. In sepsis, complement component C3 is rapidly consumed, which delays the physiologic age-dependent increase observed in healthy foals.31 Neutrophil dysfunction, characterized by decreased neutrophilic phagocytocytic ability and killing capacity via oxidative burst activity is reported in septic neonates.29-31
Pelger-Huet anomaly of neutrophils is an inherited disorder described in Arabian horses, characterized by a reduced number of nuclear segmentation and bilobed dumbbell-shaped nuclei, making mature neutrophils resemble band forms.55,56 Immunophenotyping of Pelger-Huet neutrophils of horses revealed normal expression of myeloid-specific markers and adhesion molecules, and evaluation of phagocytosis and oxidative burst via flow cytometry revealed normal neutrophil function. Despite the morphologic anomaly revealed during the evaluation of blood smears, the functions of Pelger-Huet neutrophils are virtually normal, and clinical signs of immunodeficiency are not observed; however, their inflexible nuclei restrict their ability to emigrate from blood vessels.53
Immunologic Disorders Leading to Immune-Mediated Injury
Inappropriate or exaggerated immunologic responses can directly injure bodily tissues. The clinical manifestation of immune-mediated injury depends on the nature and sites of tissue injury. Immune-mediated tissue injury can be from humoral or cell-mediated immune responses, or a combination of both. Immune-mediated tissue or organ damage, often referred to as hypersensitivity reaction, has been classified according to the nature of the immunologic pathway involved. The basic framework of disease mechanisms involving the adaptive immunity proposed by Gell and Coombs consisted of four types of immunopathologic reactions: type I (also referred to as immediate hypersensitivity) encompassing those mediated by IgE; type II encompassing cytotoxic mechanisms mediated by antibodies, generally of the IgG and IgM isotypes; type III encompassing those mediated by immune complexes, which mostly involve IgG and IgM isotypes; and type IV, which are delayed reactions mediated by sensitized T cells. The four types of hypersensitivity reactions are discussed in more detail below. Comprehensive reviews of hypersensitivity disorders may be found elsewhere.2,3,57,58
Immediate or type I hypersensitivity reactions are mediated by antibodies of the IgE isotype; hence they manifest after a primary exposure (sensitization) to antigen or allergen and subsequent B-cell differentiation, antibody production, and memory development. These reactions typically occur within 15 minutes to a few hours after the time of exposure to the antigen or allergen, depending on the need for metabolic processing of the triggering molecule. Binding of the antigen or allergen to at least two IgE molecules bound to the surface of mast cells via high-affinity IgE receptors (Fc epsilon RI) leads to degranulation of mast cells and release of mediators such as histamine, serotonin, leukotrienes, prostaglandins, various chemokines, and cytokines. These inflammatory mediators cause vasodilation, edema, smooth muscle contraction, and chemotaxis; this immediate inflammatory reaction occurs locally and potentially systemically. Of clinical significance, localized reactions may propagate to systemic reactions known as anaphylaxis. In horses, anaphylaxis may manifest with fever, hypotension, bronchoconstriction, dyspnea, tachypnea, coughing, sweating, piloerection, diarrhea, and abdominal pain, as the respiratory and gastrointestinal (GI) tracts are the shock organs.
Antibody-mediated hypersensitivities reactions include (1) reactions to self cell-associated antigenic epitopes, referred to as type II hypersensitivity, and (2) reactions involving soluble antigen-antibody (Ag-Ab) complexes, referred to as type III hypersensitivity. Antibodies of the IgG, IgM, and IgA isotypes can trigger severe inflammation via opsonization for enhanced phagocytic function and complement activation with deposition of C3b and iC3b that lead to destruction of antigen-bearing cells. In addition, Igs also mark the cell target for ADCC by NK cells. Activation of C5a potently recruits neutrophils that release proteolytic enzymes (including acid phosphatases, acid hydrolytic enzymes, cathepsins, elastase, collagenase) and oxidative agents with further tissue destruction. Target cells may be circulating or be part of tissues. In type II hypersensitivity, the binding of Ig to antigenic epitopes on cells that are fixed in a tissue or on tissue structural elements, such as basement membrane, results in damage to the respective tissue. For instance, direct antibody binding to self-antigens of the keratinocyte desmosome are responsible for damage that results in acantholysis, blistering, and desquamation in the pemphigus complex. If the antigen-bearing cell is circulating (e.g., erythrocyte, platelet, neutrophil), the cell may be destroyed in circulation but more typically is removed by the mononuclear phagocyte system. Examples of hypersensitivity to antigenic epitopes on circulating cells include alloimmune reactions (such as neonatal isoerythrolysis, neonatal alloimmune thrombocytopenia, and neonatal allogeneic neutropenia) and drug hypersensitivities (such as penicillin-induced hemolytic anemia). In the case of type III hypersensitivity, antibodies bind to soluble antigens (e.g., pathogens) that are not cell associated, resulting in Ag-Ab complexes that deposit in a variety of vascular beds, including lymph nodes, kidneys, joints, and skin.59 Tissues in the vicinity of the precipitated Ag-Ab complexes are also damaged by the ensuing immune reaction. An important condition characterized by hypersensitivity reaction to soluble antigens in horses is purpura hemorrhagica.
Delayed hypersensitivity reactions, also referred to as type IV hypersensitivity, consist of exaggerated cell-mediated immune responses within 24 to 48 hours from the time of exposure to the antigen. This condition also requires previous sensitization to the antigen, with a previous exposure that leads to APC processing and presentation of the antigens to T cells in the context of MHC and the cytokine milieu. The cytokine milieu, dictated by the APCs, leads to the differentiation of Ag-specific T cells into specific subsets of Th cells or CTLs and development of memory. Classically, the sensitization phase of cell- mediated hypersensitivity involves antigen processing and presentation in the context of MHC accompanied by IL-12 as co-stimulatory signal by the APCs, leading to differentiation of the T cells into Th1 cells. Upon reexposure to the same antigen, APCs activate the antigen-specific Th1 cells that
Features of Immunodeficiency Disorders Affecting the Adaptive Immunity of Horses
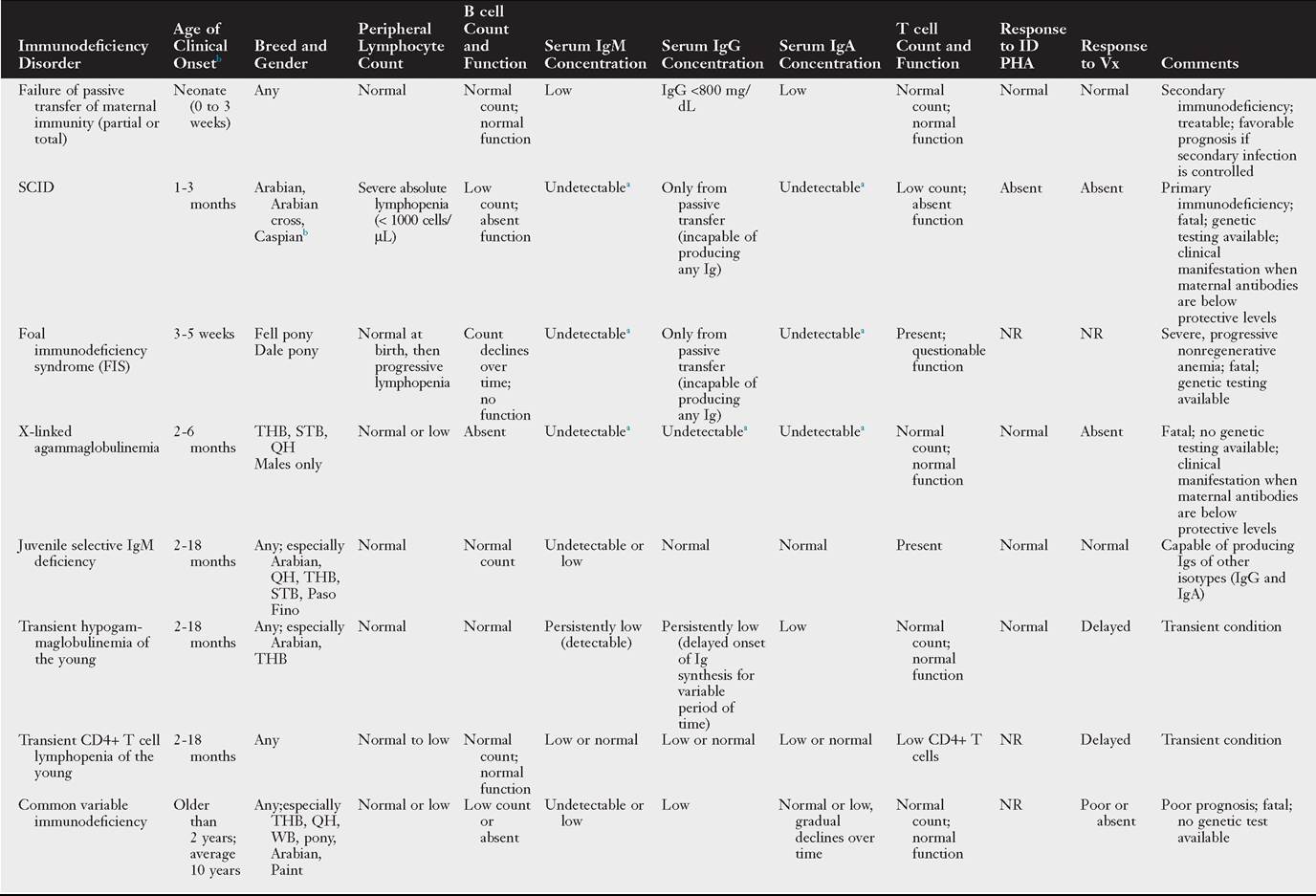
Onset of signs listed in this table is in face of adequate colostral transfer of maternal immunity, except for partial or total failure of passive transfer of maternal immunity.
aValues depend on concentration of passively transferred maternal antibodies and age.
bReport of clinical and postmortem findings of SCID, but negative for SCID genetic testing.
ID PHA, Intradermal phytohemagglutinin; Ig, immumoglobulin; NR, not reported; QH, Quarter Horse; SCID, severe combined immune deficiency; STB, Standardbred; THB, Thoroughbred; Vx, vaccine; WB, warmblood.
proliferate and produce large amounts of Thl cytokines (IFN-γ, IL-2, tumor necrosis factor [TNF]-α, and TNF-β). This cascade of cytokines activates macrophages and recruits nonspecific circulating leukocytes, including monocytes and lymphocytes, to infiltrate the site; there is amplification of the inflammatory reaction and development of epithelioid macrophages and multinucleated giant cells, leading to a chronic inflammatory reaction that mediates tissue damage or granuloma formation. If during sensitization the cytokine milieu by APCs includes IL-4 and IL-5, the T cells differentiate into Th2 cells. Upon reexposure, the sensitized Th2 cells produce large amounts of the Th2 cytokines (IL-4 and IL-5), leading to IgE production and nonspecific eosinophilic activation. If the sensitized T cells differentiate into CTLs, upon reexposure, the sensitized CTLs proliferate and mediate cytotoxicity and tissue damage. Examples of DTH in horses include (1) contact dermatitis to heavy metals and aniline dyes found in tack, bedding, soaps, and blankets; (2) granulomatous diseases; and (3) hypersensitivity to insect bite. Table 53.4 summarizes some disorders caused by immune-mediated injury in the horse; detailed discussion of these and other conditions can be found elsewhere.