Transmissible Fungal Infections
Transmissible fungal infections include those that are directly transmissible between individuals, for instance, some species of dermatophytes. Indirectly transmissible fungal infections include examples such as the water-borne chytrids Batrachochytrium Bendrobatidis and B.
Salamandrivorans. However, most pathogenic fungi are non-transmissible and are acquired from environmental sources. Exposure to a transmissible fungal infection can lead to the host becoming a carrier that harbours the infection. Carriage may lead to subclinical infection where no clinical signs of infection are manifested or clinical infection whereupon direct or indirect costs of exposure occur (Garner et al. 2009). Much of the terminology used in the wider epidemiological literature refers to the transmissible infections, because transmissible pathogens such as Plasmodium spp., Mycobacterium spp. and HIV have historically contributed the largest burden of disease and have been the focus of epidemiological modelling and efforts to control disease. Therefore, terms that refer to epidemiological characteristics of transmissible infections are relevant to fungal pathogens that are spread from host to host. Commonly used terms include the following:Attack Rate This is generally defined as the proportion of individuals who are exposed to an infectious agent and become clinically ill.
Primary/Secondary Cases For transmissible fungal infections such as Candida sp., spreading through hospital wards or Microsporum canis spreading through a population of cats, the host whom introduces the infection into a susceptible population is called the primary case, and the individuals whom are subsequently infected are called secondary cases. From these cases, the reproductive rate can be defined which describes the potential for the infection to spread through a population. The basic reproductive rate, R0, is a critical quantity to understand as it determines the probability that a new infection entering a population will continue to spread.
Defined as ‘the average number of secondary infections generated from an infected host in an entirely susceptible population', R0 predicts three possible situations following disease introduction into a population:• R0 < 1 ) the fungal pathogen will go extinct.
• R0 = 1 ) the fungal pathogen will become endemic.
• R0 > 1 ) an outbreak of the mycosis will occur.
If immunity to a fungal infection builds up within a population over time, then R0 will decay to less than 1 (for a population with an R0 of 2, then this will be the time point when roughly half the population have become immune). Therefore, determination of R0 for transmissible fungi is necessary in order to calculate the proportion of the population that needs to be vaccinated or subjected to treatment, in order to control the onwards spread of infection. As fungal vaccines and other therapies are currently being developed and deployed, this is an increasingly important area of epidemiological interest.
Index Case/Site This is the first case, or region, to be identified during an outbreak and is usually the focus of intense research as it is from the rate that secondary cases are generated and the proportion of cases that develop clinical symptoms that initial epidemiological models are formulated.
Incubation Period and Latent Period The incubation period is the time extending from when the individual is first infected through to the time that they develop symptoms of disease. This can be calculated from clinical observations or within an experimental setting; for example, Doddington et al. (2013) experimentally infected groups of Mallorcan midwife toads with two isolates of the chytrid B. dendrobatidis in order to calculate isolate-specific death rates, with which to parameterise epidemiological models aimed at assessing the probability of this species survival on the island following the introduction of the pathogen (Walker et al.
2008).The latent period is usually defined as the period of time from infection to onset of infectiousness, i.e. the stage at which secondary cases start to be generated. The latent period will vary between individuals and host species depending on the rate of growth of the fungus and time to generation of infectious inocula. However, for mycoses, latent period is often used to describe the period between the time from infection and the time to when clinical symptoms occur, so should really be termed the ‘incubation period' as described above. This usage of the term ‘latent period' is generally associated with fungal diseases such as human penicilliosis (talaromycosis) or crypto- coccal meningitis, where the time from infection to disease can sometimes be measured in decades. For both Talaromyces (previously Penicillium) marneffei and Cryptococcus neoformans, pulmonary infection can be quiescent and asymptomatic until the patient becomes immunosuppressed (either through advancing age or other causes such as HIV infection), whereupon clinical signs ensue (Shelburne et al. 2005; Walker et al. 2008).
Infectious Period This is defined as the time from onset of infectiousness to cessation of infectiousness (usually via death or recovery), i.e. the duration of infectiousness. For commensal fungal infections that are probably acquired early in life, such as Candida albicans or Pneumocystis jirovecii, the infectious period may well be lifelong for a subset of infected individuals.
Incidence and Prevalence Incidence is defined as the number of individuals whom are infected by a fungal infection during a defined period of time, often a year, divided by the total population under study. Incidence is often expressed as ?per 100,000 individuals. ‘Prevalence of infection' relates to the number of individuals who have a specific fungal infection at a specific time point divided by the total population. If the average daily incidence I of a fungal disease has a duration of D days, then the average prevalence P will be P = I x D.
In other words, ‘prevalence is the product of incidence and duration'. Prevalence of infection is often also used in conjunction with measurements of intensity of infection, as this describes the severity of infection and likelihood of severe disease and mortality. For instance, ‘Vredenburg's 10,000 zoospore rule' has found wide usage as a term that describes the average zoospore load that an amphibian host can tolerate before succumbing to chytridiomycosis (Vredenburg et al. 2010), although this value is non-absolute and varies amongst and between species (Clare et al. 2016).
Threshold Density This is the minimum population density of hosts at which transmission of an infectious fungus can be sustained with an R0 > 1.
Epidemic/Pandemic and Endemic These terms were coined by the ancient Greek physician Hippocrates in the Hippocratic corpus Of the Epidemics written around 400 B.C.E. and relate to the occurrence of diseases that were ‘normal' such as coughs and colds and ‘abnormal' such as plagues. The modern definition of epidemic has changed somewhat to ‘The occurrence in a community or a region of cases of an illness (or an outbreak) with a frequency clearly in excess of normal expectancy' (Heymann 2014). The word ‘epidemic' is therefore often linked to the concept of emerging infectious diseases as these, by definition, diseases that are in excess of normal expectancy (measured by incidence and/or prevalence of infection). Fungal pathogens that are characteristically associated with a specific region or host species are termed endemic mycoses. These mycoses are pathogenic fungi that occupy a specific niche within the environment and therefore are characterised by having spatially constrained distributions. Well-known examples are Coccidioides immitis/ C. posadasi (Fisher et al. 2001, 2002) which are associated with xeric, alkaline soils of the New World and T. marneffei which is constrained to the wet tropical region of Southeast Asia (Vanittanakom et al.
2006). Both of these species (alongside several others such as H. capsulatum and Blastomyces dermatitidis) exhibit temperature dimorphism whereupon they can grow as mycelia at environmental temperatures, but can undergo a dimorphic transition at 37 °C to grow as yeasts or spherules and cause disseminated, sometimes lethal, infection (Sil and Andrianopoulos 2015).Epizootic/Panzootic and Enzootic These terms are specific to infectious fungal pathogens that occur in a nonhuman animal population that are either wild (such as the epizootics and panzootics that are described for amphibian chytridiomycosis or Aspergillus sydowii infections of coral) (Chap. 14) or captive (such as epizootic lymphangitis in horses and mules caused by the dimorphic fungus H. capsulatum var. farciminosum) (Chap. 5). ‘Panzootic’ is the nonhuman animal counterpart to the word ‘pandemic’, and ‘enzootic’ is the nonhuman animal counterpart to the word ‘endemic’. In the context of emerging fungal infections, it is important to recognise that an epizootic mycosis can change to become an established enzootic mycosis over time as host-pathogen dynamics establish a stable equilibrium (Briggs et al. 2010). Such epizootic and enzootic dynamics have been found to occur across varied ecological systems and amphibian communities that have been affected by the introduction of B. dendrobatidis (Fisher et al. 2009; Briggs et al. 2010; Retallick et al. 2004).
Zoonoses and Zoonotic Zoonoses are infections of animals that can be transmitted to humans and vice versa. A meta-analysis of 1415 pathogens that are infectious to humans suggested that 307 were caused by species of fungi. For all classes of pathogens, Taylor et al. (2001) showed that the majority were zoonotic comprising 868 (61%); of these, 116 (38%) were considered to be zoonotic fungal infections of humans (Taylor et al. 2001; Fisher et al. 2001). However, closer inspection of the classifications shows that the authors have listed many fungal species that are actually environmental opportunistic fungal infections (or sapronoses) (Chap.
1). For instance, the genus Aspergillus, Alternaria, Acremonium, Bipolaris, Curvularia, Mucor, Paecilomyces and Rhizopus amongst many others are misclassified by Taylor et al. (2001) as ‘zoonoses’ as they are well understood by experts on fungal diseases to be acquired from their saprophytic environment. From the point of view of a mycologist, the term ‘zoonosis’ needs to be used in a more specific context to indicate animal-to- human (or human-to-animal) transmission. These ‘true’ zoonoses are relatively rare and even here there are grey areas. For instance, Cryptococcus and Coccidioides species are able to grow on nonliving substrates such as lignin (Cryptococcus) and/or soil (Coccidioides). However, both species are also able to metabolise nitrogen-rich substrates such as pigeon faeces (C. neoformans) and small-mammal corpses (Coccidioides), and these associations amplify infectious inocula, therefore increasing the risk of human infection. Arguably, these species can therefore be classified as zoonoses owing to their demonstrated association with nonhuman animals as well as human infection. True fungal zoonoses are actually rather rare and information is generally incomplete. For candidate zoonoses such as T. marneffei that infects bamboo rats in Southeast Asia, the link to human infection has never been definitively established, although the molecular epidemiology shows that human and bamboo-rat infections are identical on a molecular basis (Gugnani et al. 2004; Henk et al. 2012). Similar arguments have been made suggesting that mosquitoes can vector their microsporidian parasites such as T. hominis (Mathis et al. 2005) into immunosuppressed humans and they therefore constitute zoonotic reservoirs of these fungi. The best examples of fungal zoonoses lie within the dermatophytes such as Microsporum and Trichophyton species, where animal-to-human infection occurs frequently from companion animals (such as dogs, cats, rabbits or small rodents) or cattle (Fehr 2015) (Chap. 3). More recently, human sporotrichosis has emerged as a growing problem in Brazil, where virulent feline sporotrichosis caused by Sporothrix brasiliensis is now known to be a true zoonosis that accounts for a growing burden of human disease as infections of Sporothrix brasiliensis that have invaded urban feral cat populations spill over to infect humans (Rodrigues et al. 2014; Fehr 2015) (Chap. 10).Pathogenicity and Virulence Casadevall and Pirofski (2003) argued in a key review that both the host and their infecting microorganism contribute to disease (pathogenesis). In their article, they postulate that the amount of damage incurred by the host varies according to the relative input of pathogen and host factors, known as the ‘damage-response’ framework. Broadly, fungal pathogenicity can be defined as ‘the costs accrued through the interaction between an infectious fungus and its host' and will contain both host and pathogen components. For instance, a strong (but damaging) host immunity that is incurred through chronic stimulation of inflammatory cells will incur a cost to the host by causing disease (such as is caused by Candida albicans) (Brown et al. 2014). Conversely, direct destruction of host tissue by the fungal infection through lytic processes driven by virulence effectors, such as that caused by proteinase secretion by B. dendrobatidis (Joneson et al. 2011) or Candidalysin effector secretion by C. albicans (Moyes et al. 2016), will also incur costs to the host and cause disease. The term virulence is often (mis)used interchangeably with pathogenicity, when in fact its usage should be more refined and specific. Virulence can be understood as ‘the relative capacity of a microorganism to cause damage in a host'. The word ‘relative’ is used because virulence is often measured in comparison with other variants of the fungal species in question. Virulence is often genetically encoded, so studies commonly focus on defining specific virulence factors that are then shown to cause a specific virulence trait. For instance, in Cryptococcus gattii infection, hypervirulence in humans was shown to occur in isolates that were associated with the well-described ‘Vancouver Island’ outbreak in the late 1990s (Chap. 12). The hypervirulent isolates were genetically different from a related genotype of C. gattii that occurred in the region. These isolates were shown to have well-defined virulence traits, namely, a dramatic ability to tubularise their mitochondria in order to replicate rapidly within macrophages of the mammalian immune system that was linked to distinct genomic differences between lineages of this pathogenic fungus (Ma et al. 2009; Farrer et al. 2015).
Fungal Emerging Infectious Diseases Emerging infectious diseases (EIDs) are those infections that are increasing in either their incidence, geographic and/or host range or virulence (Daszak et al. 2000; Jones et al. 2008) and are found to occur within all groups of living organisms. Until the 1990s, EIDs caused by fungi were generally held to be a feature of plant-infecting fungi, where they have widely challenged food security ever since humans developed agricultural systems (Gurr et al. 2011). However, fungal EIDs are now recognised to occur in animals, and the frequency of their identification appears to be on the increase both spatially, temporally and in terms of their virulence (Fisher et al. 2012). The drivers that underpin this emergence of fungal disease are debated; however, it is clear that worldwide networks of trade contribute to infectious spillover into novel naive hosts as infectious inocula are moved worldwide—increased transcontinental transmission likely accounts for many of the current panzootics of fungal disease that we are witnessing (Fisher and Garner 2007) from endemic centres of infection (Fig. 2.1). Notorious fungal EIDs of animals include amphibian chytridiomycosis caused by B. dendrobatidis and B. Salamandrivorans (Martel et al. 2013) (Chap. 14), cryptococcosis caused by C. gattii lineage VGII in the Pacific Northwest of the USA (Chap. 12), bat white nose syndrome caused by Pseudogymnoascus (Blehert et al. 2009) (Chap. 13) and cat-transmitted sporotrichosis caused by Sporothrix brasiliensis (Rodrigues et al. 2014) (Chap. 10) (Fig. 2.2).
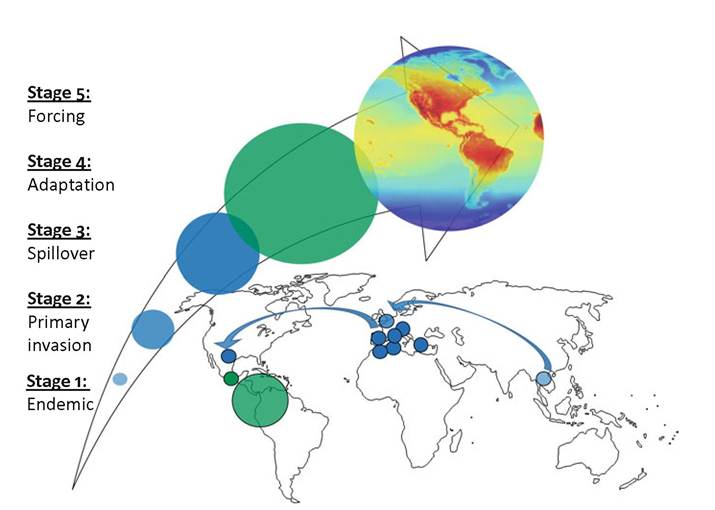
Fig. 2.1 Stages describing the emergence of a pandemic or panzootic fungal infection. In this scenario, an endemic mycosis (stage 1) invades a novel host(s) through a primary invasion event (stage 2). Subsequent spread through anthropogenic trade and/or other mechanisms leads to subsequent spillover events and establishment in new hosts and/or ecosystems (stage 3). Adaptation occurs leading to refinement of the host/pathogen interaction and survival/transmission in new hosts/ecosystems (stage 4). In stage 5, large-scale events such as environmental change may lead to further amplification of disease
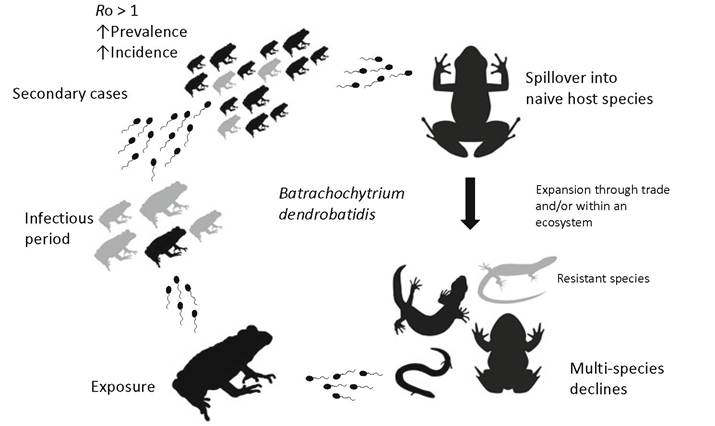
Fig. 2.2 General epidemiological features associated with an outbreak and panzootic of amphibian Chytridiomycosis
2.3
More on the topic Transmissible Fungal Infections:
- Introduction
- Transmissible Fungal Infections
- Seyedmousavi S. et al. (eds). Emerging and Epizootic Fungal Infections in Animals. Springer International Publishing,2018. - 406 p, 2018
- Abstract
- Epidemiology of Histoplasmosis
- Epidemic and Epizootic Expansions
- Index
- Abstract
- FungaL Diseases
- Infectious Ocular Diseases