Age and Functional Changes to the Female Tract
Only a general account of the important age and functional changes is presented in this chapter, which glosses over the many species differences that affect all aspects but particularly the timing and duration of events.
Age and Cyclic Changes
The juvenile reproductive organs are disproportionately small. At birth the ovaries provide no evidence of their future endocrine role, which is not established until shortly before puberty, when ripening follicles and the corpora lutea that replace them produce the hormones that stimulate the growth, tissue differentiation, and activity of the reproductive tract as well as the manifestation of female behavior. In contrast, the gametogenic or exocrine function was established in the young fetus with the migration of primordial germ cells into the ovary. These immigrant cells proliferate rapidly to produce a population of perhaps 3 million at its maximum, but this number soon begins to be progressively reduced in a process that continues to puberty and beyond. Only a few hundred thousand generally survive at birth and, because no later accession to their number is possible, the later release of female gametes is much more stingy than that of male gametes. Each surviving oocyte is initially surrounded by a single layer of flattened epithelial (granulosa) cells to form the structure known as the primordial follicle. Most primordial follicles remain in arrested development or undergo atresia, but some transform into primary follicles that are distinguished by the enlargement of the oocyte and its enclosure within a covering of granulosa cells that have assumed a cuboidal conformation.
Growth of reproductive organs is initially isometric, keeping pace with general somatic growth. After puberty the actions of ovarian hormones, cumulative over the first few cycles, bring about a rapid enlargement and a better differentiation of the component tissues.
Follicles in all stages of development may now be found within the ovaries together with corpora lutea and replacement scars (Fig. 5.56).There is a continuous slow growth of many follicles within the adult ovaries. In the ovary of anestrous animals, the follicles grow to the early antral stage (Fig. 5.57A) but then degenerate. The onset of the breeding season is heralded by a more rapid development of a few, which are chosen from this larger population through a yet poorly understood mechanism. These favored follicles enlarge at an exponential rate under the influence of FSH of the pituitary. Their growth is explained by the proliferation of granulosa and theca cells and the accumulation of follicular fluid. This fluid increasingly distends a central vesicle (antrum) into which the ovum projects, raised on a mound of cells (cumulus oophorus) and enclosed within a cellular covering (corona radiata). The follicle is bounded by a two-layered capsule (theca interna and externa) differentiated from the surrounding stroma (Fig. 5.56). As each follicle grows it shifts toward the surface of the ovary, where it forms an increasingly salient projection. The granulosa cells of the ripening follicle produce estrogen, and it is the peak level of production of this hormone that induces both the behavioral pattern and the structural changes that characterize the animal in heat.
» TABLE 5.1
Some Specific Parameters in Reproduction
| Species | Puberty (mo) | Cycle Length (days; range) | Duration of Estms | Ovulation | Pregnancy Duration (days) |
| Dog | 6-9 | ≥90 | 9 days | 3 days after Ihe beginning of estrus | 62 |
| Cat | 6-9 | Variable | 7-W days | 24 h after coitus | 63-65 |
| IIorse | 20 | 21 (19-22) | 5-6 days | 1—2 days before the end of estrus | 330 |
| CaltIe | 6-18 | 21 (18-24) | 18 h | 10—12 h after the end of estrus | 280 |
| Sheep | 6-12 | 17(16-18) | 24-36 h | 30-36 h after the beginning of estras | 150 |
| Goat | 4-8 | 21 | 24-36h | 30-36 h after the beginning of estrus | 150 |
| Pig | 5-10 | 21 (19-21) | 48-72 h | 35—45 h after the beginning of estrus | 114 |
Estrogen has an epitheliotropic effect most evident in promoting proliferation of the vaginal epithelium and simple lengthening of the uterine glands.
It also causes edema and hyperemia of the tissues of the reproductive tract; edema may produce a visible swelling of the vulva, but congestion of the endometrium may lead in some species (notably the bitch) to the appearance of blood in the external discharge. It also enhances the irritability of the myometrium that is detectable through the uterus, including the cervix, which becomes more responsive to manipulation.Ovulation occurs late in estrus or shortly after its termination and is stimulated by LH, also of pituitary origin. Ovulation is spontaneous in most species, but in some, including the cat, the mechanical stimulus of coitus is necessary to set in train the events that culminate in follicular rupture (Table 5.1). Once shed into the peritoneal cavity, an ovum is soon gathered into the expanded end of the uterine tube. Because the ovum is nonmotile, the most likely mechanisms are the production of a current in the suspending fluid by the ciliary beat of the tubal epithelium and grasping movements of the muscular fimbriae, which are closely applied to the surface of the ovary at this time. Both mechanisms would be assisted by the surface irregularity provided by adherent corona cells.
The space within the vacated follicle fills with blood when rupture has been attended by considerable hemorrhage, but any clot is soon replaced by proliferation of the surviving granulosa and internal theca cells to form a solid body, the corpus luteum (Fig. 5.57B). This structure grows rapidly and may soon equal in size the follicle that it replaces. It produces progesterone, the hormone that continues the preparation of the uterus for the reception of the embryo and for the maintenance of pregnancy. In animals that become pregnant the corpus luteum survives well into or throughout pregnancy (according to species), but it regresses quite rapidly in cycles that are infertile (Fig. 5.57C). Responsibility for its regression rests with a luteolytic hormone (prostaglandin) produced by the "empty" uterus.
The effects of progesterone reinforce those produced by previous exposure to estrogen and stimulate further growth of the uterine glands, which now become branched, tortuous, and more active, secreting the so-called uterine milk that nourishes the embryo before implantation. Progesterone also dampens the activity of the myometrium.The transport of ova within the tube is achieved by the combination of ciliary and muscular activity. If mating has occurred the ova rendezvous with the sperm within the ampulla. Although sperm may reach this site within a few minutes of coitus, a longer sojourn within the female tract is required before they become capable of fertilization. According to species, semen is initially deposited within the vagina or the cervix, where it forms a coagulum from which some sperm soon emerge. Even when the semen is deposited in the vagina, churning movements soon bring some sperm into contact with the cervical mucus, which protects and guides the sperm on their upward path. Even so, the movement of sperm would be slow if they depended on their own puny efforts; transport is mainly effected by muscular contractions, evoked by prostaglandin within the semen, and by oxytocin reflexly released into the bloodstream at coitus. Only 1% or 2% of the many millions within an ejaculate deposited within the vagina succeed in passing the cervical barrier. The uterotubal junction, the next major impediment, is successively negotiated by even fewer sperm (and these necessarily of normal motility). In species in which intrauterine deposition of sperm takes place, the uterotubal junction is the first barrier. Movement within the tube is more erratic because the muscular contractions are ill coordinated. In most species sperm remain fertile for a day or two after coitus, and many apparently find temporary refuge in cervical glands and other niches. Fertilization activates the ovum, and cleavage begins within a short time. Its later fate is considered in the following section.
The Course of Pregnancy
The evolution of the gravid uterus affects its size, position, form, and relations that become increasingly evident with advancing pregnancy. The principles effecting change in size are more or less the same in all animals, but the other aspects vary among species and are best considered separately for each (see the appropriate later chapters). The increase in size may ultimately be as much as 100-fold (as in the cow), but the greater part of this change is represented by the contents of the uterus, which comprise the fetal membranes and fluids in addition to the conceptus(es). The more modest growth of the organ involves all its components. The endometrium remains hyperemic and edematous, and the myometrium enlarges owing to a vast increase in the size of individual muscle cells. Despite this hypertrophy, the uterine wall is unable to keep pace with the growth of the contents, and it stretches markedly — so much so that in rats and other species of similar size it becomes transparent. The broad ligaments share in the increase and come to contain large amounts of muscle. The arteries enlarge greatly as it becomes necessary to satisfy an everincreasing demand for blood. Activity of the cervical glands continually renews the mucous plug that seals the cervical canal.
Implantation involves reaction from the apposed epithelial layers of the blastocyst and endometrium, and in some species considerable erosion of maternal tissue occurs during attachment (see later). This erosion occurs mainly in species in which the blastocyst remains small before implantation and either seeks out a nidus (nest) in a cleft of the endometrium or burrows into its substance. The blastocysts of domestic species grow considerably before implantation and remain centrally within the lumen of uterus. In domestic ungulates the implantation is probably delayed significantly longer than the 2 weeks after coitus suggested for many other mammals.
Implantation and the initial development of the fetal membranes concludes the preembryonic period, the first of the three periods into which development is conventionally divided.
Its principal features may be summarized as follows: the intrauterine migration and eventual settlement of the blastocyst, and its rapid transformation from a spherical to a threadlike form in many species (which include the ruminants and pig but not the horse).The second or embryonic period includes the establishment of a fully functional placenta, the differentiation of the various tissues and organ systems, and the initiation of various functions, most notably an embryonic circulation. By the end of this period the external conformation is sufficiently developed to identify the major taxon—order and, perhaps, family—to which the embryo belongs, though not yet the particular species.
The remaining part of the intrauterine development is somewhat arbitrarily and imprecisely assigned to a third, or fetal, period.* Organogenesis continues throughout the fetal period and, for many organs, well into postnatal life, but the changes that now bring the different systems into the levels of structural and functional competence necessary for survival after birth are less dramatic than those that took place earlier. The rapid growth, which is the foremost characteristic of the fetal period, continues into postnatal life without significant interruption around the time of birth (Fig. 5.65).
The early transformations and the complexities of organogenesis provide ample opportunity for death or malformation in the first two periods. This is probably true for all mammals, although data are most reliably available for the human and the pig. Some losses and abnormalities are due to intrinsic defects of the conceptus, some to an unreceptive state of the uterus, and some to exposure of the mother to any of a variety of environmental insults. It is known, for example, that chromosomal abnormality of structure or number is demonstrable in about 10% of clinically detectable human pregnancies, including spontaneous early abortions, and it is believed to be even more common in conceptuses lost at earlier stages, before there was awareness of pregnancy. In contrast, chromosomal abnormality is identified in a much smaller proportion, perhaps 0.5%, of human infants delivered at term. Although the fertilization rate in pigs is high, possibly exceeding 95% in some herds, it has been estimated that only 60% or so of conceptuses come to term. Most deaths occur within the first 40 days (of a gestation period of 114 days), but because conceptuses lost at an early stage are generally resorbed and leave no trace, the figure must be interpreted with caution. The rates of fertilization and delivery at term for other species vary too much from herd to herd and stud to stud to be conveniently summarized.
The environmental insults that may affect development include ionizing radiation, viral infections, inorganic and organic chemicals, including some that are constituents of plants (e.g., clover, soya and certain other legumes, Veratrum californicum) potentially present in pasture or other feedstuffs. Many of these agents are better known from their effects in the laboratory than in the field, and although some are lethal, others are more likely to produce abnormalities that are survivable, if only for a time. Such agents (teratogens) are most likely to produce abnormality when exposure occurs during the embryonic period, when so many complicated and critically timed procedures are under way.
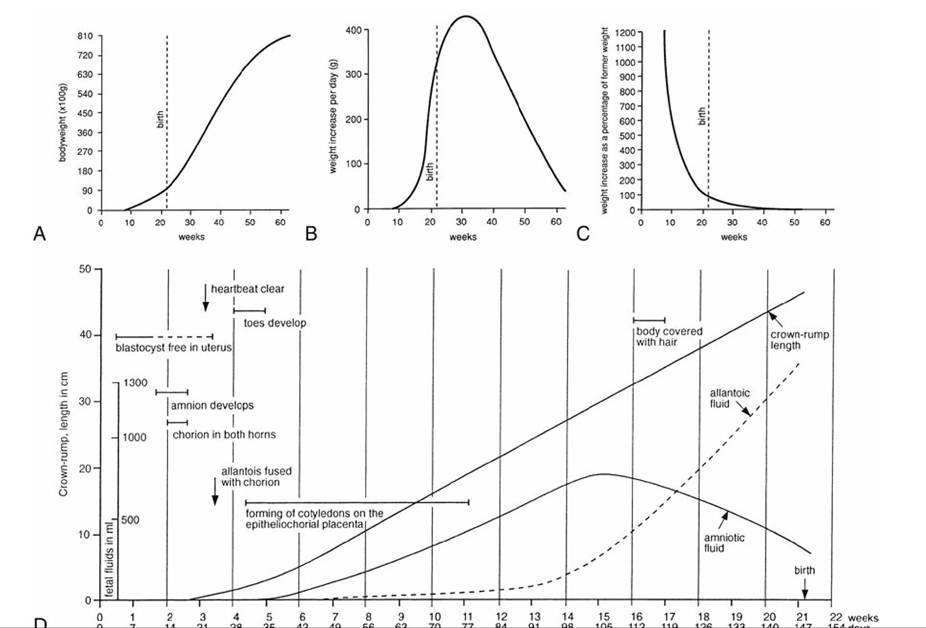
FIG. 5.65 The growth of lambs. (A), (B), and (C) record the growth in weight of lambs during fetal and early postnatal life. (D) Schematic summary of metrical and other features of the development of the fetal lamb and its adnexa.
Infectious agents such as bovine viral diarrhea (BVD), hog cholera (swine fever) (HCV), border disease virus (BDV), and human rubella and cytomegalovirus can cause fetal death, resulting in abortion or stillbirth and producing defects of brain and eye especially or growth retardation in young born to mothers infected in early pregnancy. Fetuses infected at a later stage with BVD or HCV become immunotolerant to these viruses and may be born apparently healthy. Because they are persistently infected, they represent a real danger to other livestock on the farm.
Fetal Membranes and Placentation
We have insufficient space to describe the formation of the embryonic or fetal membranes but include diagrams (Fig. 5.66) as reminders of the principal points. The definitive gross arrangement is shown for the dog (Fig. 5.67A), horse (Fig. 5.67B), and ruminant (Fig. 5.67D and E) conceptuses. The fetal membranes and the endometrium participate in the formation of the placenta, an organ that facilitates physiologic exchange and hormone production. A provisional placenta, furnished by a vascularized yolk sac, may provide a useful organ of exchange in early pregnancy. This omphaloplacenta is important in the first third or so of equine pregnancy (Fig. 5.68), but in most species the chorioallantoic placenta, the definitive placenta of eutherian mammals, becomes competent at a relatively earlier stage. In the definitive arrangement, the chorion, intimately associated with the endometrium, is vascularized by vessels that reach it by following the allantoic outgrowth from the hindgut. The stalk of the allantois (urachus), the accompanying vessels that become the umbilical arteries and veins, and the ensheathing connective tissue (the fetal variety known as Wharton's jelly) constitute the umbilical cord, which persists as the communication between fetus and placenta until ruptured in the course of birth or shortly thereafter.
The chorioallantoic placenta takes many forms that may be classified in several complementary ways. The first system refers to the gross distribution of the chorionic villi, minute outgrowths of the chorionic surface that engage with depressions of the endometrial surface to provide the areas of exchange. The horse and the pig have a diffuse placenta in that the villi are spread in small clumps (microcotyledons) over virtually the entire surface of the chorion (Fig. 5.67B and C). In ruminants the villi develop in scattered patches or cotyledons opposite the endometrial caruncles; each cotyledon and associated caruncle forms a separate unit or placentome, which collectively constitute a cotyledonary placenta (Fig. 5.67D and E). In the dog and the cat the villi develop in a band of chorion that encircles the trunk of the embryo, forming a zonary placenta* (Fig. 5.67A). In the fourth and last type, discoidal placenta, which commonly occurs in primates and rodents but not in domestic species, the villi are concentrated in one large patch (Fig. 5.69).
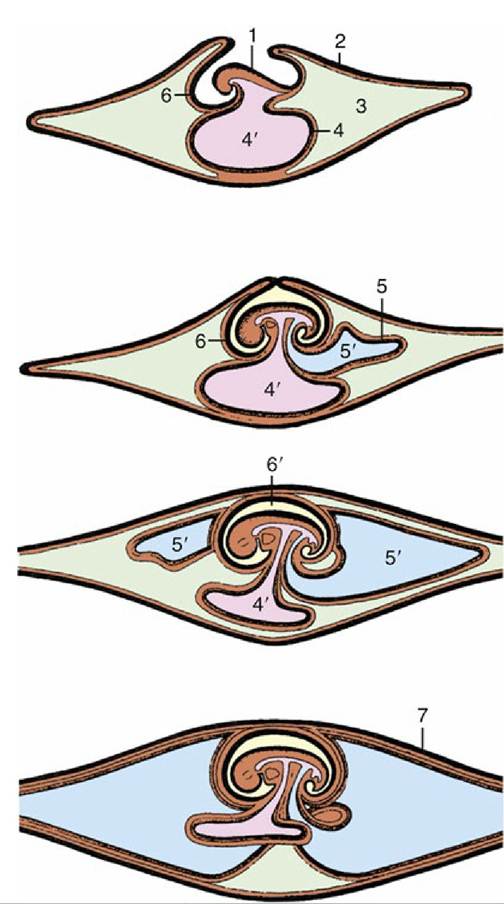
FIG. 5.66 Schematic representations of the formation of extraembryonic membranes. 1, Embryo; 2, chorion; 3, extraembryonic celom; 4, yolk sac; 4', yolk sac cavity; 5, allantois; 5', allantoic cavity; 6, amnion; 6', amniotic cavity; 7, chorioallantois.
The second system refers to the tissue layers that separate the fetal and maternal bloodstreams. Initially, six layers are present: chorionic capillary endothelium, connective tissue, epithelium, endometrial epithelium, connective tissue, and capillary endothelium. The tissue barrier at the areas of exchange is always later reduced, sometimes only by the closer approach of the two sets of capillaries but often by tissue loss. In theory, the six layers persist in the epitheliochorial placenta seen in the mare and sow. They are reduced to four, by the loss of the endometrial epithelium and connective tissue, in the endotheliochorial placenta seen in dogs and cats (Fig. 5.70), and they suffer the ultimate reduction to one layer, embryonic endothelium, in the hemoendothelial placenta of bats. Ruminants were long described as having a syndesmochorial placenta, in which only the uterine epithelium had been lost; modern studies discount this loss, and it is now believed that these animals also have epitheliochorial placentas.
The third system refers to the loss of maternal tissue that occurs at birth. In some species the fetal and maternal layers part cleanly, no maternal tissue is shed, and the description nondeciduate is appropriate. When implantation is interstitial, considerable maternal loss may be expected; the human placenta is of this deciduate type. Minor loss of uterine tissue occurs in an intermediate semideciduate type found in ruminants (Table 5.2).
The histologic system appears to define different degrees but offers incomplete explanations of variations of placental permeability. The barrier may not be exactly as implied by the description. Moreover, the placenta evolves and changes in structure during pregnancy, and significant regional differences may exist side by side. Freely diffusible molecules cross from one circulation to the other, and in this respect the human hemochorial placenta certainly allows more rapid passage than the "thicker" epitheliochorial placenta of the larger domestic species. The transport of larger molecules depends on other factors, including specialized unidirectional mechanisms.
Differences in the barrier to the passage of immunoglobulin G (IgG) are of particular veterinary significance. In some species a mechanism exists for the transfer of maternal antibodies to the fetus to confer some immediate protection on the newborn. This prenatally acquired immunity is denied to offspring of species (including horses and farm animals) with epitheliochorial placentas, and their neonates rely on colostrum, the milk first produced, as the source of antibodies for temporary protection.
Fetal antigens, present in plasma or borne by blood cells, may leak into the maternal bloodstream with potentially damaging consequences. The classic illustration is furnished by the hemolytic disease of human infants (erythroblastosis fetalis). This condition develops in a second or later child confronted by antibody produced by a Rhesus-negative mother in reaction to a previous Rhesuspositive child. Antibody production by the mother develops so slowly that the child (usually the first) provoking the response generally escapes serious harm. Similar conditions occur in other species, including horses and pigs, but damage to their offspring can be prevented by denying them access to colostrum, which contains the relevant antibody.
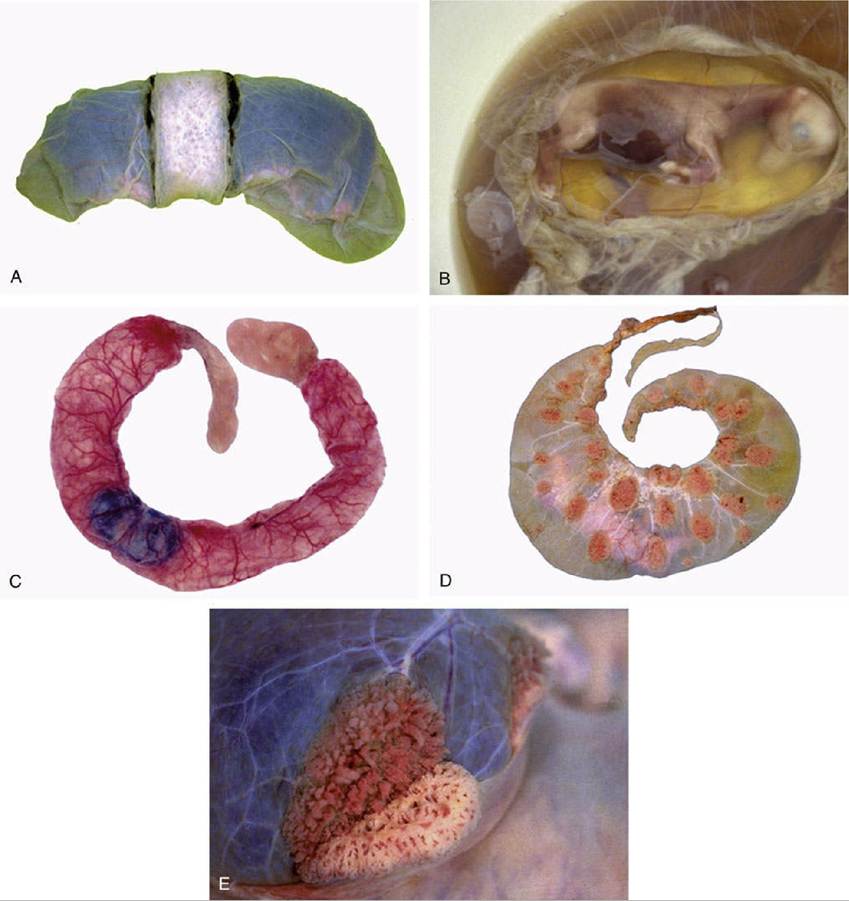
FIG. 5.67 Placentation in a number of species. (A) The most intense interaction between the fetal membranes and the endometrium in domesticated species is found in carnivores with the zonary placenta. (B) The placenta of the horse fetus is not very complex. The villi do not penetrate deep into the endometrium. (C) A similar situation exists in the pig. (D) The placenta of ruminants is a cotyledonary placenta with many placentomes (cow). (E) The partial separation of the maternal and fetal part of the placentome is demonstrated.
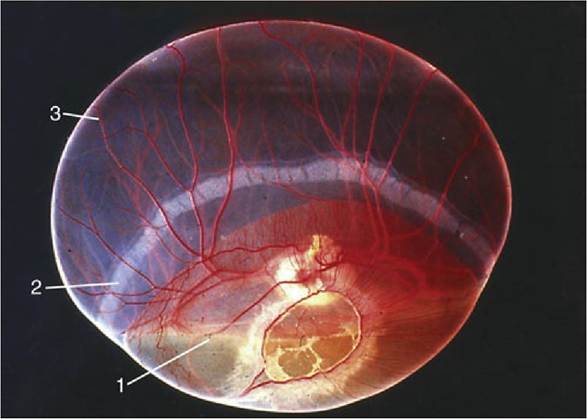
FIG. 5.68 Young conceptus (horse). 1, Yolk sac; 2, chorionic girdle; 3, allantochorion.

FIG. 5.69 Discoidal placenta of a rat.
The endocrine functions of the placenta are both complex and lacking in uniformity among species, even between those that are closely related. The horse is of unusual interest in this context because equine chorionic gonadotropin is produced from structures unique to Equidae: the endometrial cups (Fig. 5.71 and p. 564). Elevated concentrations of steroid hormones in blood and urine provide the basis for diagnosis, including the do-it-yourself tests for pregnancy in women. Tests of comparable reliability, simplicity, and economy are not yet available for domestic species, for which reliance is still largely placed on clinical procedures, including ultrasonography.
The placenta is one (though not the only) source of other hormones relevant to pregnancy. Lactogen, a hormone related to growth hormone, acts with other hormones to develop the mammary glands for the approaching lactation, while relaxin, which is also secreted by the corpus luteum, helps prepare the reproductive tract and pelvic parietes for parturition (p. 564); later, acting in synergy with oxytocin, the latter stimulates the expulsive activity of the myometrium.
Prostaglandin is a product not of the placenta but of the endometrium of the "empty" uterus, and its production is delayed for 2 weeks (or so) after the corpus luteum first forms. It leaves the uterus in the uterine vein, and in some species, including ruminants, it reaches the ipsilateral ovary after countercurrent transfer to the ovarian artery. In others, for example, the horse, in which contact between artery and vein is less close, it reaches the ovaries via the general circulation. Within the ovary, prostaglandin promotes Iuteolysis (regression of the corpus luteum) with consequent decline and eventual cessation of progesterone secretion and release. The production of prostaglandin is stimulated by oxytocin, but in pregnancy the conceptus produces a factor that blocks endometrial receptivity to oxytocin, thus indirectly protecting the corpus luteum, whose integrity is now required.
Before we conclude the account of the fetal membranes and placenta, brief attention may be given to the fluids contained within the amniotic and allantoic cavities. These fluids, dramatically released at the time of birth—the "breaking of the waters"-actually have rather important functions to perform throughout gestation, and at certain stages they account for a very considerable fraction of the total content of the gravid uterus. The fluid within the amniotic cavity immediately surrounds and supports the embryo or fetus, cushioning it against compression and abdominal trauma. This protection is most required by the young embryo, whose skeleton is still largely unformed and whose external covering—hardly yet to be called skin—is delicate and vulnerable to trauma. Later, when these structures are better developed, the amniotic fluid tends to be reduced in amount (see Fig. 5.65). At its maximum, it measures about 3 to 5 L in cattle and perhaps a little more in horses; in pigs it varies around 100 mL, and it is about 10 to 30 mL in dogs and cats. The amniotic fluid has a brisk turnover with roughly matched production and resorption in the short term. In early stages, the fluid is a dialysate from vessels of the embryonic skin and amnion; later, once rupture of the urogenital membrane has opened a passage from the bladder, it consists largely of urine, and as more is added, the fluid already present is reduced by being swallowed. Deficient and excessive amounts of this fluid (oligohydramnios and polyhydramnios, respectively) are possible complications of pregnancy, the former often indicating anomalous development of the kidneys, the latter potentially open to correction by the addition of a "sweetener" to encourage deglutition. This fluid is not normally a significant contributor to that present in the respiratory passages of the fetus and newborn, contrary to some suggestions. Being slightly mucoid, amniotic fluid has additional value as a lubricant of the birth canal at parturition.
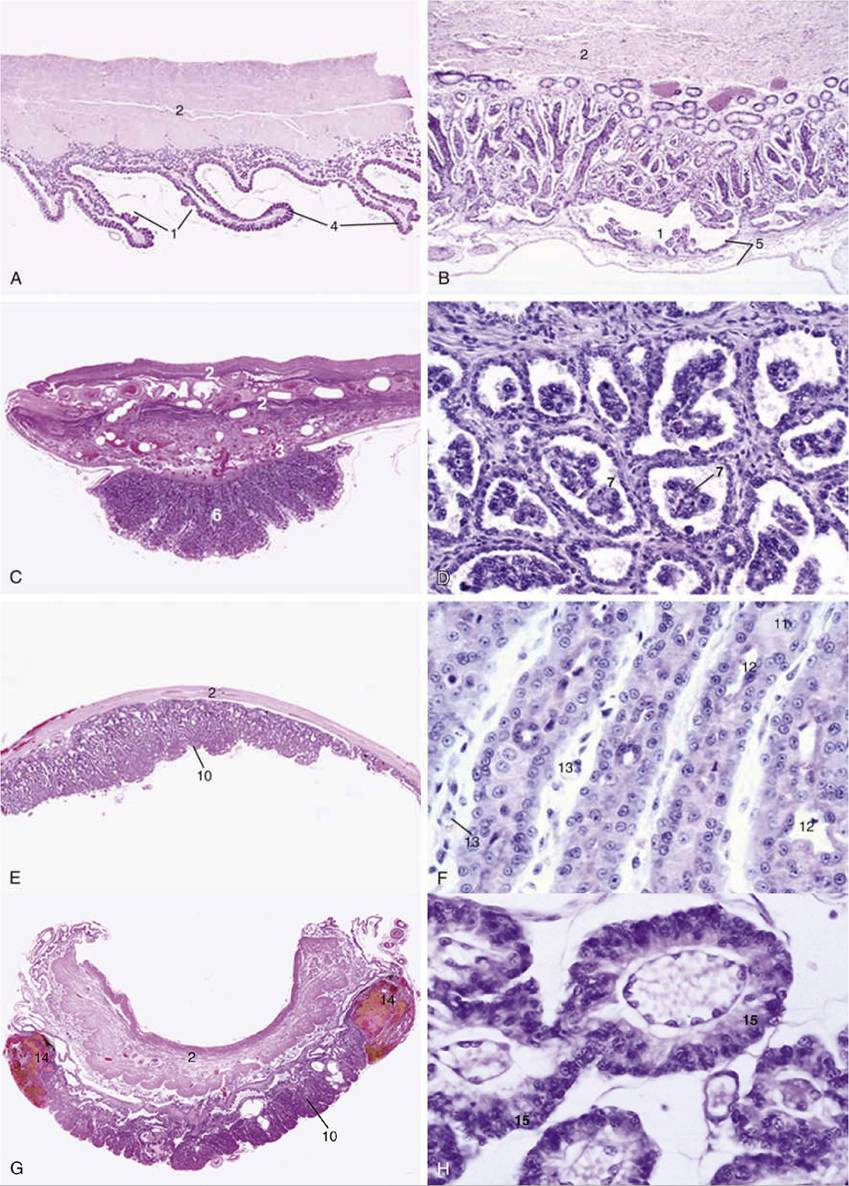
FIG. 5.70 (A) to (H) Placental histology; (H) for label descriptions. (A) Diffuse folded villous placenta (pig) (4?). (B) Diffuse villous placenta (horse) (28?). (C) and (D) Cotyledonary villous placenta (cow) (4?; 140?).
(E) and (F) Zonary labyrinthine placenta (cat) (4? and 279?). (G) and (H) Zonary labyrinthine placenta (dog) (4? and 279?). 1, areola; 2, myometrium; 3, endometrial glands; 4, primary fold; 5, allantochorion; 6, placentome; 7, trophoblastic giant cells; 8, uterine septum; 9, chorionic villi; 10, placental labyrinth; 11, decidual cell; 12, maternal capillaries; 13, fetal capillaries; 14, marginal hematomas; 15, trophoblast cells.
» TABLE 5.2
Placental Classification
| Carnivores | Horse | Ruminants | Pig | |
| Gross form | Zonary and labyrinthine | Diffuse | Cotyledonary | Diffuse and folded |
| IIistologic type | Endotheliochorial | Epitheliochorial | Epitheliocliorial | Epitheliochorial |
| Separation | Sernidecidnate | Nondeciduate | Semideciduate | Nondeciduate |
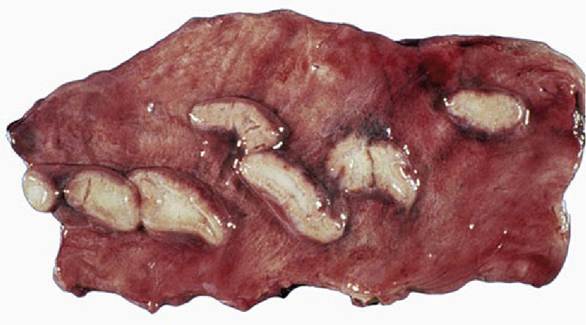
FIG. 5.71 Endometrial cups (mare) during early pregnancy. These cups are responsible for the production of the pregnant mare's serum gonadotropin (PMSG).
The allantoic cavity is large in all domestic species, but the human allantois fails to expand and is soon reduced to a negligible vestige. It is possibly the consequent lack of medical interest that explains the relative paucity of information concerning the formation, turnover, and role of allantoic fluid. The allantoic cavity does, of course, receive urine through the urachus before the urethral route is established, and this process helps maintain the osmotic pressure of the fetal plasma at a level that prevents fluid loss to the maternal bloodstream. A second function may be to maintain sufficient radial pressure to hold the chorion tight against the endometrium in those species in which the placental attachment is less firm. There is rather more allantoic than amniotic fluid in the large species, and amounts are about the same in dogs and cats. Although there is about 100 mL in the pig at midterm, the quantity is reduced to very little at full term. However, the quantities are rather variable in all species.