BASIC PLAN AND DEVELOPMENT
Although the forelimbs and hindlimbs are not homologous, they have a similar organization and segmentation with a remarkably close correspondence of analogous parts. Each first appears as a bud that grows out from the ventrolateral surface of the body of the young embryo at a level corresponding to the origin of the nerves by which it will later be supplied.
The bud of the forelimb appears before that of the hindlimb, and its development maintains this advantage for some time— indeed, until after birth in puppies and other animals born in a rather immature state. These animals initially confine their locomotor activities to dragging themselves, using the forelimbs only, toward their dam’s teats.When first formed, a limb bud consists of a mass of mesenchyme, the loose embryonic connective tissue, within an ectodermal covering. The ectoderm becomes the epidermis, including its derivatives; the mesenchyme differentiates to form skeletal tissues, muscles and tendons, fasciae, and blood vessels. Thus it is only the limb nerves that invade from outside; all other structures develop in situ. The limb bud lengthens, and its free distal part expands to form a flattened hand (foot) plate while the more proximal part acquires a more columnar form. Thickenings corresponding to the digital rays soon appear in the plate and are accentuated when the intervening tissues are reduced. The details of this development naturally vary with the species, for it is only some that retain the primitive pentadactyl (fivedigit) pattern and only a few that show a complete separation of digits. It is interesting to note that five digits appear in most species; when evolution has reduced the complement to fewer, the adult condition is usually attained by fetal regression of some digits. Creases formed in the proximal part of the bud soon allow recognition of segments corresponding to the arm and forearm (or thigh and leg) regions of the adult.
The first indication of the future limb skeleton is provided by an axial condensation of the mesoderm to produce a denser core. In the early stages of development (but not always later) a definite proximodistal gradient of differentiation occurs. This establishes and then maintains the girdle elements in advance of those of the arm or thigh, and the latter in advance of more distal parts.
In the next stage of development the mesoderm is locally transformed to create a series of cartilaginous models in the pattern of the adult bones. These precursors soon come to resemble the final forms in broad outline; they remain ensheathed by thin coverings of the unmodified mesoderm, now appropriately known as perichondrium. Dense mesoderm also remains between the cartilages where the joints will develop.
The cartilage models grow mainly by interstitial growth, in which each part expands more or less uniformly to maintain the general form. The next stage involves the replacement of the cartilage by bone tis- sue—not its transformation into bone, a distinction that deserves to be emphasized. The process does not occur identically or synchronously in different bones, and the remarks that follow concern that hypothetical concept, the “typical long bone.”
The initial ossification involves two processes. In one, the perichondrium around the middle of the shaft lays bone down on the cartilage. This process of bone formation is known as intramembranous ossification because it occurs within the connective tissue membrane. Its details must be sought in textbooks of histology. A tubular bony sheath, the periosteal collar, is thus formed about the center of the shaft; it is gradually extended toward each extremity (Figure 2-43). In the
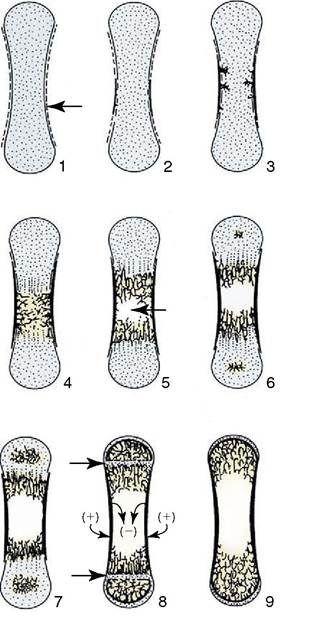
Figure 2-43 Development of a long bone, schematic. 1, Cartilage model with perichondral membrane (arrow);2, intra- membranous ossification of diaphysis; 3, 4, endochondral (primary) ossification of diaphysis, replaces cartilage; 5, beginning of medullary cavity (arrow); 6, epiphysial ossification centers appear; 7, endochondral (secondary) ossification of epiphyses; 8, narrow epiphysial cartilages (arrows) separate the diaphysis from epiphyses: these and the articular cartilages are all that remain of the cartilage model (1); note circumferential growth of diaphysis by removal (-) and addition (+) of compact bone; 9, mature bone consisting of articular cartilage, spongy bone, and compact bone; the epiphysial cartilages have disappeared.
other process, the cartilage of the center of the shaft shows aging or degenerative changes; its cells hypertrophy, come to occupy enlarged lacunae (spaces) in the matrix, and then die, while the matrix becomes impregnated with calcium salts. This central patch of dead cartilage is now invaded by a connective tissue sprout that pushes in from the periosteum (as the perichondrium is now more appropriately known in the region of the collar). The progress of this sprout, which is rather cellular and well vascularized, is facilitated by the spongy texture given to the dead cartilage by the enlarged lacunae. Some of the cells that are carried inward have the capacity to engulf and remove calcified matrix, others have the capacity to lay bone down on the surviving framework, while a third group are precursors of marrow cells. The processes of construction and destruction continue in parallel and transform the whole middle portion of the shaft into a parcel of bone known as the primary or diaphysial center of ossification.
Later (much later in some species and mainly after birth in ourselves), similar sprouts from the perichondrium invade the centers of the two extremities; they establish secondary or epiphysial centers of ossification. The secondary centers are not preceded by the formation of any equivalent to the periosteal collar of the shaft. The general stage of development of the long bone at this time is shown in Figure 2-43/5. This reveals that the original cartilage now survives only as two plates, the epiphysial or growth cartilages, that intervene between the primary and secondary centers. These have a special significance since they are responsible for the growth in length of the bone. They are clearly polarized; cell division and matrix expansion are confined to the epiphysial aspect while degeneration, calcification, and replacement occur at the central or diaphysial side (Figure 2-44). The replacement adds continuously to the length of the diaphysis while the growth of the cartilage continues to shift the epiphyses away from this.
The two processes are balanced until finally growth fails to keep pace with replacement. The plate thins and ultimately is quite destroyed. The epiphysis and diaphysis have now fused as one, and further longitudinal growth is impossible. Neither the rates of growth nor the times of final disappearance are necessarily the same in the two growth cartilages of a long bone. Meanwhile, however, the bone has also been increasing in its girth, which is the result of further lamellae being laid in succession on the existing bone within the periosteal sheath. Some of the larger projections on long bones develop from independent centers of ossification and remain separated from the shaft by cartilage growth plates while growth continues. The projections distinguished in this way are known as apophyses.Little reflection is necessary before deciding that bone growth must be more complicated than this. The form established by the original model would not be maintained by continuous accretion. A simultaneous process of destruction must exist, especially to maintain the shape of the metaphyses (the regions of the shaft adjoining the growth cartilages), to keep surface features in the same relationship to each other, and to establish and then enlarge the medullary cavity. Although we have no space to elaborate on this statement, one point can be made: bone grows by apposition, the deposition of new material on that previously existing. In this it differs from the periosteum, which grows interstitially as though uniformly stretched. The periosteal sheath therefore shifts relative to the underlying bone, and the consequent drag on the nutrient vessels explains the generally oblique orientation of the adult nutrient foramina. By the time of birth, skeletal development has reached very different stages in different mammalian species. In the precocious ungulates, immediately active after birth, almost all epiphyses are well established at term. This contrasts sharply with the much less mature condition of the canine and, most especially, human neonates, in which many of the secondary ossification centers have yet to appear.
The individual rate of skeletal development is affected by many factors—inherited, nutritional, and hormonal, the last covering a complex situation in which hormones of hypophysial, thyroid, adrenal, and gonadal origin are involved. It is hardly surprising that abnormality of skeletal development is common.
Figure 2-44 Equine (pony) fetus 80 days. The developing skeleton has been colored with Alcian blue and Alizarin red. The calcified parts are red (Alizarin) and the epiphyses have not begun to calcify. These cartilaginous parts are blue.
The important features of the development of joints can be discussed more briefly. The joint tissues derive from the mesoderm left between the cartilaginous primordia of the bones. Spaces that develop in this tissue coalesce to form a single synovial cavity bounded by articular cartilage and synovial membrane. The former is probably produced by delayed chondrification of the mesoderm bordering the cartilaginous models; structural differences suggest that it is not the outer shell of this model left over after completion of epiphysial ossification. The synovial membrane is a more direct transformation of the mesoderm bordering the space. The fibrous part of the capsule and periarticular ligaments develop from more peripheral mesoderm.
It is now generally agreed that the limb muscles develop within the buds. The attractive notion that portions of myotomes migrate into these buds, pulling along the appropriate nerves, has been abandoned. Certain mesenchymal cells outside the denser axial core differentiate into precursor muscle cells (myoblasts); these then increase in number through mitosis while recruitment from the mesenchyme continues. These myoblasts then form myocytes or muscle cells by a maturation in which the nuclei increase in number and migrate to the periphery of the cells. The final number of muscle cells seems, in most species, to be established before birth, perhaps well before birth.
The later growth of muscles therefore depends on an increase in the size of existing elements.The limb nerves grow in from the ventral rami of certain spinal nerves: generally C6-T2 for the forelimb and L4-S2 for the hindlimb. The segmental pattern becomes disturbed by the development of the limb plexuses in which fibers from the several ventral rami reassort before combining as the named peripheral trunks. As a consequence, all but a few very small muscles are supplied by fibers that lead from neurons in more than one spinal segment. The sensory fibers to the skin arrange themselves so that specific regions are more or less the territory of particular spinal segments. The basis for this has become more difficult to understand now that it is believed that the dermis of the limb skin develops from cells of local origin, not from cells that migrated from particular somites.
Table 2-3 lists in parallel columns the bones of the forelimb skeleton and the parts to which they give support; for comparison, columns for the corresponding bones and parts of the hindlimb (which, it will be recalled, are analogous and not homologous) are also included. A central column gives additional terms, more common in zoological than in veterinary literature, that are common to both limbs; most are not used in this text but may be encountered elsewhere.
Some entries in the first and last columns may include three terms. Those printed in plain type are the technical words used when referring to domestic animals, the terms commonly employed by veterinarians; those italicized are the corresponding words used in human anatomy; and those in brackets are the more elevated Latin terms. Probably the most surprising feature of the table is the apparent absence of vernacular terms for certain regions of animals. The situation is in fact rather better, or rather worse according to one’s point of view, than it appears. Many additional vernacular terms are restricted by custom to certain species; for example, the metacarpus of the horse is known as the cannon, but that of the dog is not. A particular difficulty is presented by the lack of handy equivalents to “paw” in description of farm animals: manus and pes are unacceptably pedantic (hence enclosed in brackets), and forefoot and hindfoot are usually (if not entirely logically) preferred; however, to the horse owner the foot generally means only the hoof and its contents. It is impossible to avoid all inconsistency.
In this book we employ the more elevated terms where it seems that vernacular equivalents might be ambiguous, risking the charge of pedantry. It is of course more sensible to use the everyday terms in conversation with laypeople.
| Table 2-3 Terms in Use for the Parts and Bones of the Limbs | ||||
| FORELIMB | Terms Common to Both Limbs | HINDLIMB | ||
| Body Part | Skeleton | bgcolor=white>SkeletonBody Part | ||
| Shoulder region, | Scapula and clavicle | Cingulum (girdle) | Os coxae (hip bone) | Pelvis |
| shoulder | Ilium Pubis Ischium | |||
| Arm, upper arm | Humerus | Stylopodium | Femur (properly os | Thigh (femur) |
| (brachium) | femoris) | |||
| Forearm | Radius and ulna | Zeugopodium | Tibia and fibula | Leg (crus) |
| (antebrachium) [Manus] | [Autopodium] | [Pes] | ||
| Carpus, wrist | Carpal bones | Basipodium | Tarsal bones | Hock, ankle (tarsus) |
| Metacarpus | Metacarpal bones | Metapodium | Metatarsal bones | Metatarsus |
| Digit, finger | Proximal, middle, and | Acropodium | Proximal, middle, and | Digit, toe |
| distal phalanges | distal phalanges | |||