THE SKELETON OF THE FORELIMB
Pectoral Girdle
The scapula, or shoulder-blade (Figure 2-45), is a flat bone that lies over the laterally compressed, craniodor- sal part of the thorax, where it is held in place by an arrangement (synsarcosis) of muscles without forming a conventional articulation with the trunk.
It is the basis of the shoulder region, a term that embraces much more than the immediate neighborhood of the shoulder joint. In ungulates, the scapula is extended dorsally by an unossified portion, the scapular cartilage (Figure 2-45, E/13), which enlarges the area for muscular attachment. The cartilage becomes increasingly calcified and thus more rigid with age.The bone is roughly triangular, though less so in the dog and cat than in the other domestic species. Its lateral surface is unequally divided by a prominent spine into supraspinous and infraspinous fossae, each occupied by the like-named muscle. The spine extends from the dorsal border almost to the articular angle and may bear a thickening for the insertion of the thoracic part of the trapezius; it is generally palpable through the skin. In all but the horse and pig, it ends in a prominent process (acromion), laterally flattened to form a hamate process in the carnivores (Figure 2-45/7') and furnished with an additional projection (suprahamate process; Figure 2-45/7") in the cat. The medial surface of the bone is largely given over to the origin of the subscapularis, which occupies a shallow fossa; a more dorsal roughened area, where the serratus ventralis attaches, extends onto the cartilage in the larger species.
The caudal border is thickened and almost straight. The thinner and sinuous cranial border is notched toward its distal end for the passage of the suprascapular nerve. The dorsal border is also generally straight and extends between cranial and caudal angles; the latter is thickened and more easily identified on palpation.
The ventral or articular angle is joined to the body of the bone by a slightly constricted neck. Its caudal part carries a shallow glenoid cavity (Figure 2-45/12) for articulation with the head of the humerus. The cavity, which is somewhat extended in the sagittal direction, faces more or less ventrally. A large muscular process, the supraglenoid tubercle, projects in front of the cavity; it gives origin to the biceps brachii.The clavicle is reduced to a fibrous intersection in the brachiocephalicus. A nubbin of bone in the dog and a slender rodlet in the cat are embedded in the intersection; their sole importance lies in the risk of misinterpretation when they are seen on radiographs.
Skeleton of the Free Appendage
The humerus (Figure 2-46) forms the skeleton of the arm. It is a long bone that lies obliquely against the ventral part of the thorax, more horizontally in the large species than in the small. It is also relatively shorter and more robust in horses and cattle than in the small ruminants and carnivores. The proximal extremity carries a large articular head (Figure 2-46/2), facing
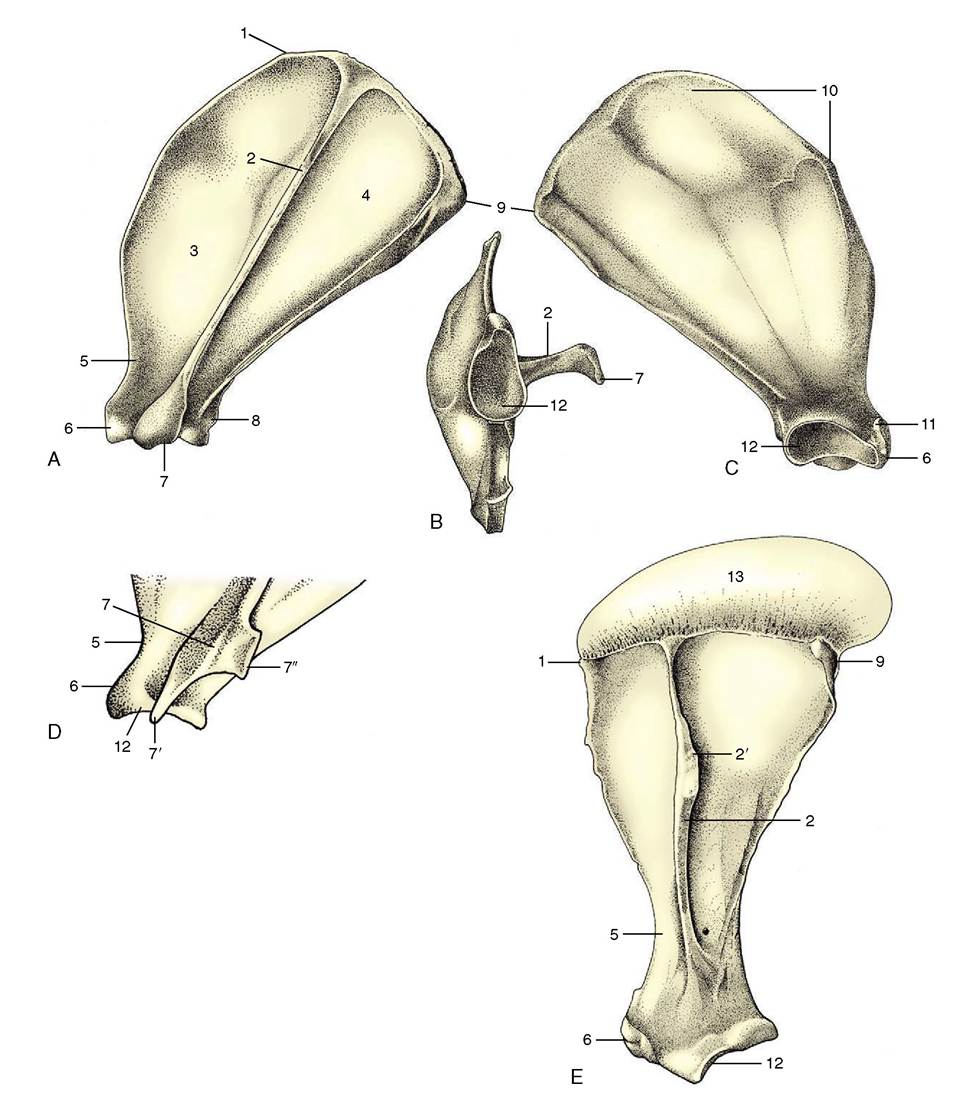
Figure 2-45 Left scapula of the dog; lateral (A), ventral (B), and medial (C) views. Distal end (D) of left feline scapula. Left equine scapula (E). 1, Cranial angle; 2, spine; 2', tuber of spine; 3, supraspinous fossa; 4, infraspinous fossa; 5, neck; 6, supraglenoid tubercle; 7, acromion; 7, 7'', hamate and suprahamate processes of acromion; 8, infraglenoid tubercle; 9, caudal angle; 10, facies serrata; 11, coracoid process; 12, glenoid cavity; 13, scapular cartilage.
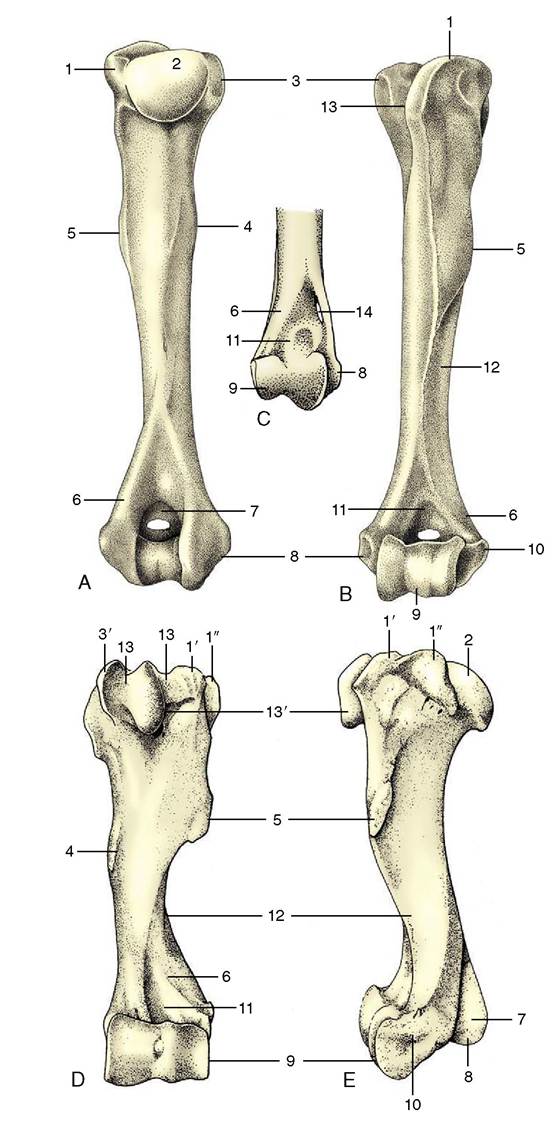
toward the glenoid cavity of the scapula and thus offset to the shaft to which it is joined by a neck. The head is shaped like a segment of a sphere and is considerably larger than the fossa with which it articulates.
Two processes, the greater (lateral) and lesser (medial) tubercles, are placed in front and to the side of the articular area.Figure 2-46 Left humerus of the dog; caudal (A) and cranial (B) views. C, Distal end of right feline humerus; cranial view. Cranial (D) and lateral (E) views of left equine humerus. 1, Greater tubercle; 1', 1", cranial and caudal parts of greater tubercle; 2, head; 3, lesser tubercle; 3', cranial part of lesser tubercle; 4, teres (major) tuberosity; 5, deltoid tuberosity; 6, lateral supracondylar crest; 7, olecranon fossa (with supratrochlear foramen in dog); 8, medial epicondyle; 9, condyle; 10, lateral epicondyle; 11, radial fossa; 12, groove for brachialis; 13,intertubercular groove; 13', intermediate tubercle; 14, supracondylar foramen.
They are separated by the intertubercular groove (Figure 2-46/75) through which the biceps tendon runs. The processes are sometimes more or less equal, as in the horse. More often the lateral one, which forms the basis of the surface feature known as the point of the shoulder, is larger; it is so in the dog. In the horse and in cattle, both tubercles are divided into cranial and caudal parts (Figure 2-46/7',7",5'); the intertubercular groove is also molded by an intermediate tubercle in the horse (Figure 2-46/75'). The medial and lateral tubercles give attachment to the muscles that brace and support the shoulder joint, substituting for collateral ligaments.
A twisted appearance is imparted to the shaft by a groove (Figure 2-46/72) that spirals over the lateral aspect and carries the brachialis and the radial nerve. Laterally, toward its upper end, the shaft carries the large, easily palpated deltoid tuberosity (Figure 2-46/5), which is joined to the greater tubercle by a prominent ridge. A less prominent, gradually subsiding ridge, the crest of the humerus, continues distally beyond the deltoid tuberosity. The medial aspect of the shaft is marked by the much less salient roughening, the teres (major) tuberosity.
The distal extremity bears an articular condyle (Figure 2-46/9) that is also set at an angle to the axis of the shaft. In large animals it engages with the radius and has the form of a trochlea. In the dog and cat it is divided into a medial area (trochlea) for the ulna and a lateral area (capitulum) for the radius. In all species the caudal part of the groove of the trochlea is continued proximally into a deep (olecranon) fossa (Figure 2-46/7) that receives the anconeal process of the ulna. Two saliences proximal to the articular surface are known as epicondyles. The medial one (Figure 2-46/5) is prominent and forms a right-angled, caudally directed projection that gives origin to the flexor muscles of the carpus and digit. The cranial aspect of the lateral epicondyle (Figure 2-46/70) gives origin to the extensor muscles of the carpus and digit. To the side, each epicondyle gives origin to the corresponding collateral ligament of the elbow joint. In the dog the floor of the olecranon fossa is perforated by a supratrochlear foramen that opens to a much shallower radial fossa on the cranial aspect of the shaft (Figure 2-46/7,77). In the cat alone, the medio- distal part of the humerus is pierced by a supracondylar foramen (Figure 2-46/74) that gives passage to the median nerve and brachial artery.
The skeleton of the forearm is provided by two bones, the radius and the ulna (Figure 2-47). In the standing position they are arranged with the ulna caudal to the radius in the upper part of the forearm but lateral in the lower part. In the primitive condition these bones articulate only at their extremities, leaving an interosseous space between their shafts; rotational movements of the human forearm bones result in turning the hand so that the palm is brought to face forward (supination) or backward (pronation). In most domestic animals the capacity for these movements is reduced or lost, and the two bones are firmly held together by ligaments or by fusion in the prone position.
When supination is possible, it consists of rotation of the upper extremity of the radius within the embrace of the ulna while the distal extremity is carried in an arc around the ulna.Clearly, no movement is possible when the bones are fused, which is the condition prevailing in ungulates and reaching its extreme in the horse, in which only the upper end of the ulna remains distinct (Figure 2-47, D/7). About 45° of supination is allowed to the dog, and somewhat more to the cat. (Rotation at the carpus contributes a substantial extra component to the movement subjectively interpreted as supination.)
The radius is a rather simple rodlike bone, usually much stronger than the ulna in ungulates, but less dominant in carnivores, in particular the cat. The proximal extremity is transversely widened, though tending to a more circular plan in carnivores, in which some supina- tory capacity remains. It articulates with the distal articular surface of the humerus and is shaped to match this. A circumferential facet (Figure 2-47, B/9) on the caudal part of the extremity articulates with the ulna and is present even when no supination is allowed. The shaft is craniocaudally compressed and slightly bowed in its length. The distal part of the cranial surface is grooved for the passage of the extensor tendons (Figure 2-47, C), whereas the caudal surface is roughened for muscular attachment. The medial border is subcutaneous and therefore palpable.
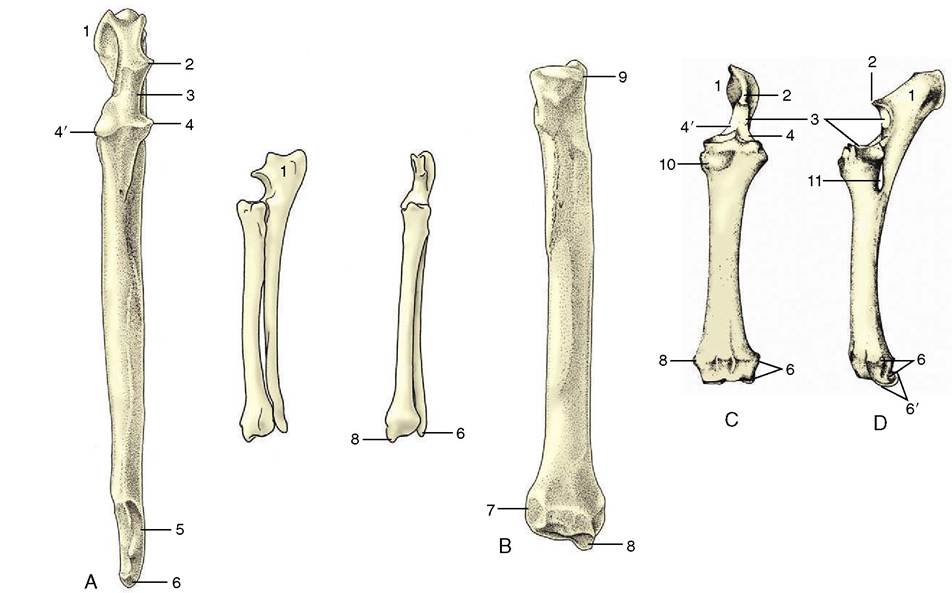
Figure 2-47 Left ulna (A) and left radius (B) of the dog. In sequence from the left: cranial view of the ulna, Craniolateral and cranial views of the radius and ulna, and caudal view of the radius alone. Cranial (C) and lateral (D) views of fused left radius and ulna of the horse. 1, Olecranon; 2, anconeal process; 3, trochlear notch; 4, 4', lateral and medial coronoid processes; 5, distal articular facet for radius; 6, lateral styloid process (with facet for the ulnar carpal bone in the dog); 6', distal end of ulna incorporated within radius; 7, articular facet for ulna; 8, medial styloid process; 9, circumferential facet; 10, radial tuberosity; 11, interosseous space.
The distal extremity of the radius is somewhat expanded.
It carries an articular surface that is concave in its cranial part and convex in its caudal part in ungulates; it has a slightly concave ovoid form in carnivores, in which some abduction, adduction, and rotation are allowed to the antebrachiocarpal joint in addition to the major movements of flexion and extension. Medial to the articulation, the radius is prolonged to form a styloid process (Figure 2-47, BZV). The corresponding lateral projection is furnished by the ulna and, in the horse, by the portion of the radius representing the incorporated ulna.The ulna has an unusual appearance, as its shaft is greatly reduced and its proximal extremity is prolonged beyond the articular surface to form the high olecranon, the point of the elbow. This process, which constitutes a very prominent landmark, gives attachment to the triceps. Distal to this, the cranial margin carries the beaklike anconeal process (Figure 2-47Z2), which fits into the olecranon fossa of the humerus, above an articular notch that engages with the humeral trochlea; yet farther from the extremity, there is a facet for the circumferential articular area of the radius. In the dog the shaft, though slender, runs the full length of the radius from which it is separated by an interosseous space that is bridged by membrane in life. The distal extremity carries a small articular facet for the radius and beyond this is continued as the lateral styloid process (Figure 2-47Z6), which makes contact with the ulnar carpal bone.
Reduction of the ulna is greatest in the horse, in which the shaft tapers to end at midforearm level (Figure 2-47, D). The distal part became incorporated within the radius in fetal life (Figure 2-47Z6'). The ruminants and pig show intermediate conditions. Of course the fusion of the ulna with the radius prohibits the movements of supination and pronation in the domestic mammals other than the dog and cat.
The short carpal bones articulate in complex fashion. The plan of the primitive carpal skeleton is uncertain, but in domestic species the bones are clearly arranged in two rows (Figure 2-48). The proximal row comprises (in mediolateral sequence) radial, intermediate, ulnar, and accessory bones; the last appears as an appendage projecting behind the carpus and is a prominent landmark in the live animal. The radial and intermediate carpals fuse in the dog and cat. The elements of the distal row are numbered from one to five (again in mediolateral sequence), although the fifth never appears as a separate bone but is either suppressed or fused with the fourth. The first is also often lacking while the second and third fuse in ruminants. The diagrams illustrate the carpal formulae in different species. Apart from the accessory carpal bone, which is probably a sesamoid by origin, a small sesamoid bone is embedded in the medial tissues of the joint of the dog. Intrinsically unimportant, it can confuse radiographic interpretation by wrongly suggesting a “chip” fracture.
Viewed as a whole, the carpus is convex from side to side on its cranial aspect and flat and very irregular caudally, although in life these irregularities are smoothed by thick ligaments. Most movement occurs at the antebrachiocarpal level, some at the intercarpal level, and virtually none at the carpometacarpal level or
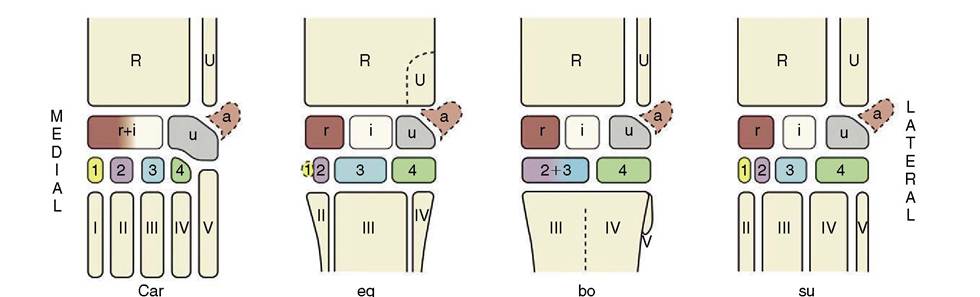
Figure 2-48 The bones of the carpal skeleton in the carnivores (Car), horse (eq), cattle (bo), and pig (su), schematic. Roman numerals identify the metacarpal bones; Arabic numerals, the distal carpal bones. R, Radius; U, ulna; a, accessory carpal bone; i, intermediate carpal bone; r radial carpal bone; u, ulnar carpal bone.
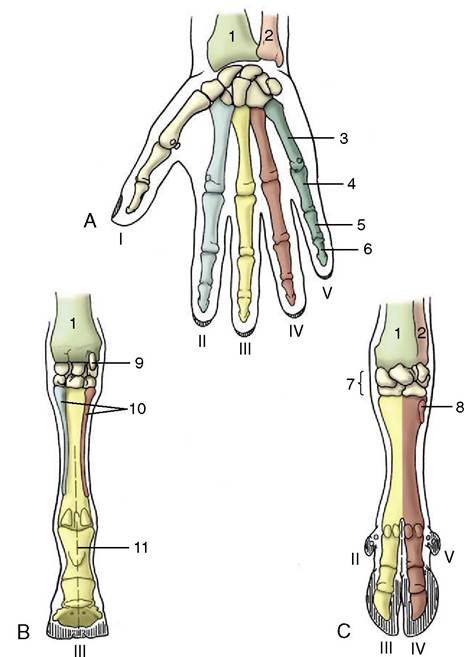
Figure 2-49 Right manus (human hand; A) of horse (B) and ruminant (C), palmar views. The Roman numerals number the rays. 1, Radius; 2, ulna; 3, metacarpal; 4, 5, 6, proximal, middle, and distal phalanges; 7, carpal bones; 8, rudimentary metacarpal V; 9, accessory carpal bone; 10, rudimentary meta- carpals II and IV (medial and lateral splint bones); 11, axis in line with ray III (mesaxonic), in C paraxonic.
between neighboring bones in a row. The combined proximal articular surface is the reciprocal of that of the radius (see earlier) and in carnivores has a convex ovoid form.
The primitive pattern for the skeleton of the mammalian manus exhibits five more or less equal rays, each consisting of a metacarpal bone and proximal, middle, and distal phalanges in line (Figure 2-49, A). This pattern has been modified in all domestic species, each of which (not excepting the pig) is to some degree specialized for fast running. Cursorial specialization involves raising the manus (and pes) from the primitive “flatfooted” (plantigrade) posture demonstrated by bears (Figure 2-50). An intermediate stage, the digitigrade posture, has been attained by dogs, which support themselves by the digits only; it culminates in the unguligrade posture attained by ruminants, pigs, and horses,
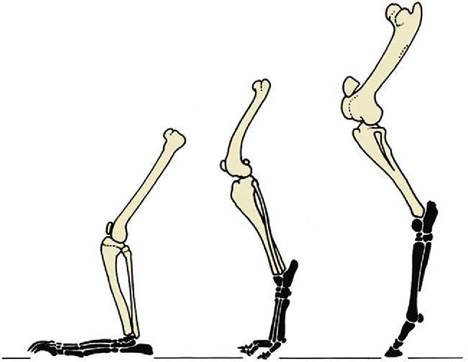
Figure 2-50 Hindlimbs of bear, dog, and horse (from left to right) illustrating plantigrade, digitigrade, and unguligrade postures.
in which only the tips of the digits, protected by hooves (ungulae), give support. The process results in the abaxial digits first losing permanent contact with the ground; a compensating development of the remaining digits enables them to carry an increased proportion of the weight. The process has not progressed very far in the dog and cat, in which only the most medial (first) digit has lost contact and is retained as a nonfunctional dewclaw (Figure 2-51). The four functional digits are broadly equal, with the axis of the manus passing between the third and fourth digits (a paraxonic position). Pigs have entirely lost the first digit; the second and fifth digits are very much reduced, although each retains a complete skeleton. In ruminants the process has gone further, and although elements of four digits are present, those of the abaxial pair are vestigial; the metacarpal bones of the functional third and fourth digits are fused in a single bone that retains evidence of its composite origin (Figure 2-49, C).
In the horse (Figure 2-49, B), only the third ray survives in functional form and its axis coincides with that of the limb; the manus is said to be mesaxonic. Remnants of the second and fourth metacarpal bones survive as the splint bones that flank the third metacarpal or cannon bone; they end in nodules, but the assumption that these incorporate greatly reduced elements of all three phalanges of the lost digits is unfounded.
The differences in the metacarpal and digital skeleton are very striking as a consequence of these changes, and the short description that follows is amplified in later chapters by details of a species-specific nature.
As the number of metacarpal bones diminishes, the relative stoutness of the surviving members of the series
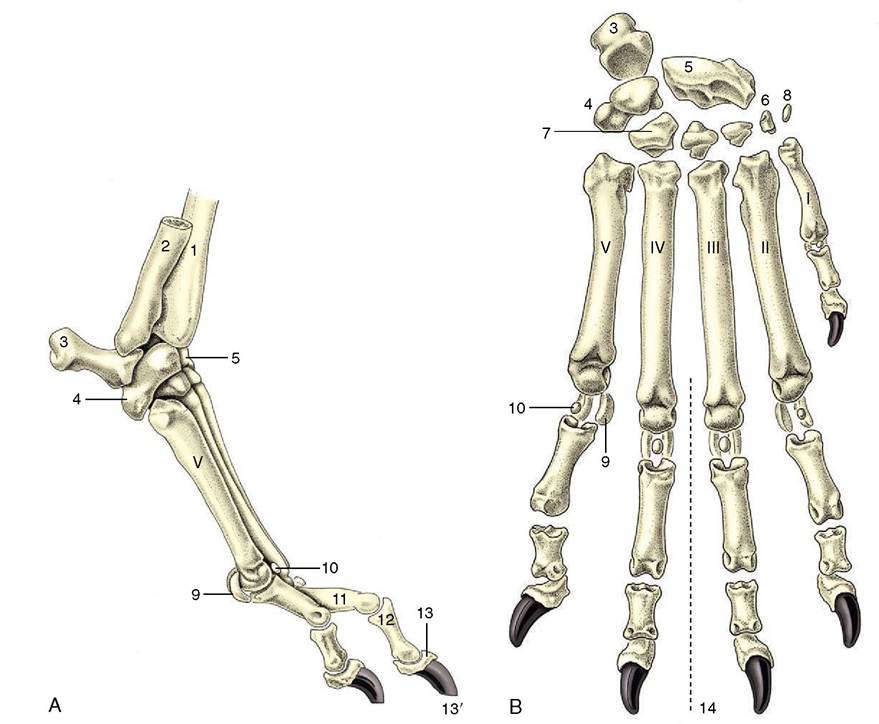
Figure 2-51 Skeleton of the right manus of the dog, lateral (A) and dorsal (B) views. The Roman numerals identify the metacarpal bones. 1, Radius; 2, ulna; 3, accessory carpal; 4, ulnar carpal; 5, radial carpal (intermedioradial in the dog); 5', intermediate carpal; 6, 7, first and fourth of the distal row of carpal bones; 8, sesamoid bone; 9, proximal sesamoid bones; 9', ridged articular surface of equine metacarpus III, articulates with proximal sesamoid bones (not shown); 10, dorsal sesamoid bone; 11, 12, 13, proximal, middle, and distal phalanges; 13', claw; 14, axis of manus.
increases. The single (third) metacarpal bone of the horse therefore has a particularly strong shaft, whereas the individual metacarpal bones of the dog are relatively much weaker. The dog’s bones are also shaped by their mutual contacts; the third and fourth bones are square in section, and the flanking second and fifth bones are triangular. Taken as a whole, the metacarpal skeleton of all species is somewhat compressed in the dorsopalmar direction. Each bone has a proximal extremity (base), a shaft, and a distal extremity (caput). The base has a flattish articular surface for the distal row of carpal bones and may, according to its position in the metacarpal series, have medial and lateral facets where it makes contact with neighbors. The distal extremity articulates with the proximal phalanx by a hemicylindrical surface with a central ridge. Various roughenings for ligamentous attachment are present at both extremities.
The proximal phalanx is a short cylindrical bone with a proximal extremity adapted to the caput of the metacarpal bone and a distal articulation in the form of a shallow trochlea. Again, the bone may be shaped by its position in the digital series.
The middle phalanx is shorter than, but basically very similar to, the first phalanx. The distal phalanx corresponds to the form of the hoof or claw in which it is wholly (hoof) or partly (claw) contained. The digital skeleton is completed by paired proximal sesamoid bones at the palmar aspect of the metacarpophalangeal joint and by a distal sesamoid bone (cartilage in the dog) at the palmar aspect of the distal interphalangeal joint. In the dog small sesamoids also exist within the extensor tendons over the dorsal aspect of the metacarpophalangeal joints.