Bones
The primary functions of the skeleton are to support the body, to provide the system of levers used in locomotion, and to protect soft parts. Therefore, biomechanical factors are most important in shaping the bones and in determining their microscopic design.
The major skeletal tissue, bone, has a secondary role in mineral homeostasis: being a reservoir for calcium, phosphate, and other ions.The Classification of Bones
Bones may be classified in various ways. A topographic classification recognizes a cranial skeleton (of the head) and a postcranial skeleton consisting of two divisions: the axial skeleton of the trunk and the appendicular skeleton of the limbs. A second classification based on ontogeny distinguishes the somatic skeleton, formed in the body wall, from the visceral skeleton, derived from the pharyngeal (branchial) arches. A third system also based on development distinguishes parts preformed in cartilage (and later largely replaced by bone) from those that ossify directly in fibrous connective tissue. This classification reflects phylogeny, in that bones that develop in membrane are homologous with dermal bones of lower vertebrates.
Bones are also classified on the basis of shape (Fig. 1.13).
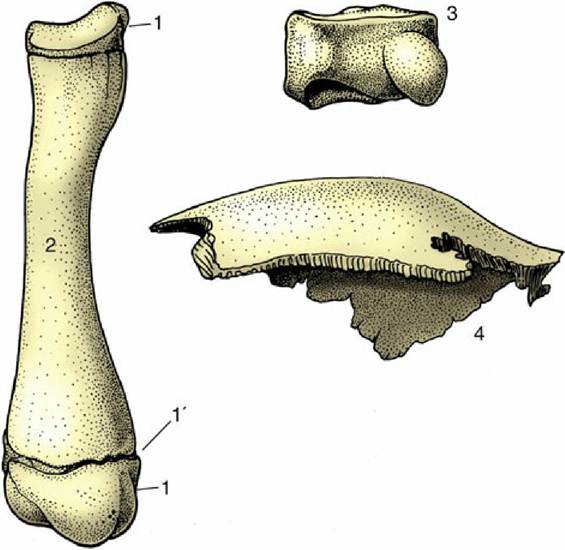
FIG. 1.13 Long (left), short (top right), and flat (bottom right) bones. 1, Proximal and distal epiphyses; 1', epiphysial cartilage; 2, diaphysis of a young dog's radius; 3, carpal bone of a horse; 4, parietal bone from the skull of a dog.
Long bones, as in the limbs, are broadly cylindrical and act as levers. They develop from at least three centers of ossification: one for the shaft (diaphysis) and one for each extremity (epiphysis) (p. 65).
Short bones have no dimension that greatly exceeds the others. Many such bones are grouped together at the carpus and tarsus to generate multiple articulations for facilitating complex movements and diminishing concussion.
The majority of short bones develop from a single center of ossification; replication of centers generally indicates that the bone represents the fusion of elements distinct in ancestral forms.Flat bones are expanded in two directions. The category includes the scapula, the bones of the pelvic girdle, and many of those of the skull. Their broad surfaces afford attachment to large muscle masses and protection to underlying soft parts.
The remaining bones are too irregular in form to be grouped in clearly defined categories. Neither flat nor irregular bones exhibit uniformity in development.
The Organization of a Long Bone
The longitudinal section of a long bone shows many features of its construction (Fig. 1.14A). The form of the bone is determined by a sheath or cortex of solid (compact) bone that is composed of thin lamellae arranged mainly in series of concentric tubes about small central canals. Each such system is known as an osteone (Fig. 1.14B). The cortex is thick toward the middle of the shaft but thins as it flares toward each end, over which it continues as a crust. The external surface is smooth except where irregularities serve as the attachment sites for muscles or ligaments; these irregularities may be raised or depressed and, in either case, permit a concentration of the attachment. These features are generally most pronounced in larger, older males. They are given a variety of descriptive names of conventional significance; most elevations are known as lines, crests, tubercles, tuberosities, or spines; most depressions are known as fossae or grooves (sulci).
The inner surface of the shaft bounds a central medullary (marrow) cavity and is rough; the irregularities are low and without apparent significance.
The extremities are occupied by cancellous or spongy bone, which forms a three-dimensional lattice of interlacing spicules, plates, and tubes of varying density.
The medullary cavity and the interstitial spaces of the spongy bone are occupied by bone marrow, which occurs in two intergrading forms.
Red bone marrow is a richly vascularized, gelatinous tissue with hematopoietic properties; it produces the red and granular white corpuscles of the blood. Although all marrow is of this type in the young animal, most is later infiltrated with fat and converted into waxy yellow marrow whose hematopoietic potential is dormant. It is the marrow in the larger spaces that first becomes inactive, followed by that of the spongy bone of the distal limb bones, until finally active marrow is confined to the proximal ends of the humerus and femur, the bones of the limb girdles, and those of the axial skeleton. The chronology of these events for domestic animals is uncertain.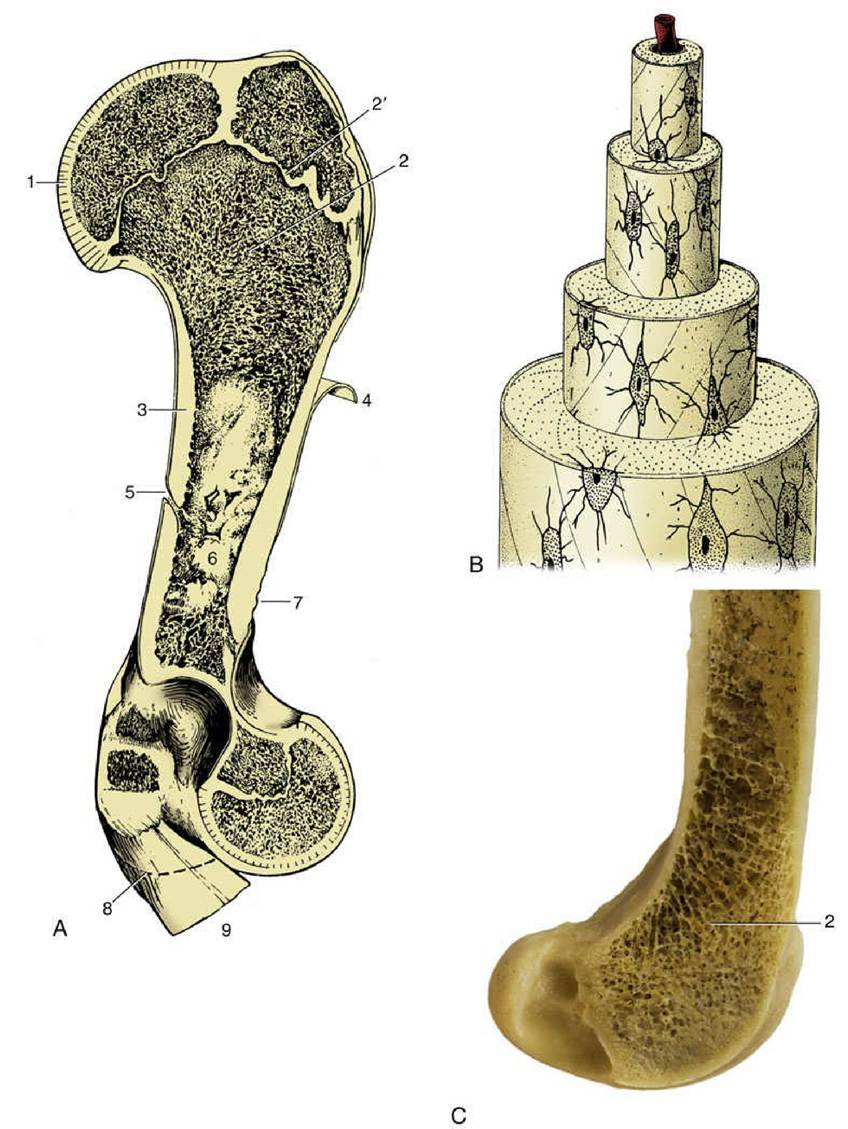
FIG. 1.14 (A) A schematic view of long bone (bovine humerus) sectioned longitudinally. (B) Osteone with central (haversian) canal. (C) Femur of dog sectioned sagitally. 1, Articular cartilage; 2, spongy bone; 2', epiphysial cartilage; 3, compact bone; 4, periosteum, partly reflected; 5, nutrient foramen; 6, medullary cavity; 7, roughened area for attachment of muscle or ligament; 8, distal extent of medial epicondyle; 9, tendons of origin of carpal and digital flexors.
The articular surfaces of bones are more extensive than the areas in contact in any position of the joint and provide a range of movement. They are smooth and clothed in hyaline articular cartilage. The cartilage is not uniform in structure; it is calcified in its deepest layer, which is firmly attached to the underlying cortex, and becomes fibrous toward the periphery, where it blends with the periosteum and joint capsule.
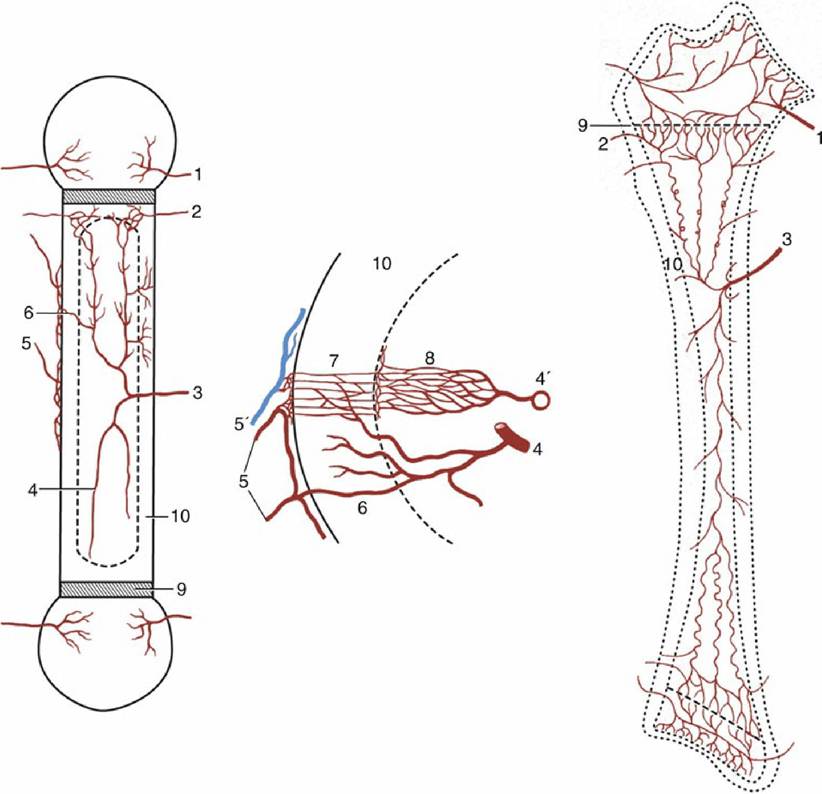
FIG. 1.15 The blood supply of a long bone, schematic. The supply of the cortex is shown (enlarged) in the center. 1, Epiphysial arteries; 2, metaphysial arteries; 3, nutrient artery; 4, 4', artery and vein of the bone marrow, respectively; 5, periosteal arteries; 5', periosteal vein; 6, anastomosis between periosteal and bone marrow arteries; 7, capillaries of the cortex; 8, sinusoids in the bone marrow; 9, growth cartilage;
10, cortex.
A tough fibrous membrane, the periosteum, ensheathes the remainder of the outer surface, from which it can be readily stripped, except where it is penetrated by tendons and ligaments to anchor in the compacta. Its appearance is rather misleading because the deeper layer is cellular and, even in adults, retains the bone-forming capacity that is exercised during development. This osteogenic function is reactivated in the healing of a fracture.
Bones have a generous blood supply, perhaps amounting to 5% to 10% of the cardiac output. Several sets of vessels exist; the so-called nutrient artery, though generally the largest single source, probably contributes less than the others in the aggregate. The nutrient artery penetrates toward the middle of the shaft in a position that is fairly constant for each bone. It is usually directed toward one end of the bone, and the foramen through which it passes may simulate an oblique fracture when depicted in radiographs. The two branches of the artery diverge and further branch within the marrow and pursue very tortuous courses, which may reduce the pressure within the vessels of the delicate marrow (Fig. 1.15). The smaller branches supply the sinusoids of the marrow tissue and also the arterioles and capillaries that permeate a system of tiny central channels (haversian canals) within the osteones of compact bone. A further supply to the cortex arises from the medullary sinusoids. Branches of the nutrient artery that reach the metaphyseal region (the part of the shaft adjacent to the epiphysis) anastomose there with branches of metaphyseal and epiphyseal vessels that enter the bone toward its extremity. The central region of this part of the shaft probably relies mainly on the nutrient artery, whereas the peripheral part relies on metaphyseal arteries. The collateral circulation is generally sufficient to allow a fractured bone to survive deprivation of part of its usual supply. One technique employed in fracture repair (intramedullary pinning) is possibly even more damaging to the vessels than the initial injury, and its success serves to emphasize the value of the anastomoses.
There is some debate about additional blood supply to the cortex from small but numerous periosteal arteries.The main drainage of the marrow is effected by large, thin-walled veins that accompany the major arteries and emerge through the nutrient, epiphyseal, and metaphyseal foramina. The capillaries within cortical tissue drain into venules within the periosteum. The normal cortical circulation is therefore centrifugal—from within outward. No lymphatic vessels are present within bone, although infections of bone may spread to the lymphatics that drain neighboring tissues.
One important difference is exhibited by the circulation in young growing bones. In them, the circulation within the epiphyses forms separate and independent compartments, because (with few exceptions) arteries do not penetrate the growth (epiphyseal) cartilage.
Nerves accompany the larger vessels, and their branches are to be found within the central canals of the osteones. Some (vasomotor) fibers pass to the vessels, some are sensory to the bone tissues (especially the periosteum), and the destination of others remains unclear. It is no longer believed that nerves exert a trophic influence on bone.
Biomechanical Aspects
It has long been the convention to explain the tubular construction of long bones by drawing the comparison with a loaded beam of some stiff, homogeneous material supported at both ends (Fig. 1.16). In this construction the tensile forces that tend to disrupt the material are concentrated toward the lower surface while the compressive forces that tend to crush and compact the material are concentrated toward the upper surface. These forces tend to neutralize each other along, and close to, the axis, and the material here is more or less redundant. It can be dispensed with or replaced by some weaker but lighter material, as in a long bone. The analogy is not exact—for a start, bone is a composite material—but it is useful as a first approach. The diagram in Fig. 1.16 shows that the lines of principal compressive and tensile stress intersect in orthogonal fashion toward the extremities of the model; the spongy architecture of a bone (Fig.
1.17) closely mimics the theoretical pattern.Compact bone is a plastic, composite material of considerable strength, capable of sustaining and recovering from considerable deformation. When it is bent, the lamellae and osteones of which compact bone is constructed first shear past each other; if the bone is bent too far, a crack appears at right angles to the line of shear and then quickly spreads to create a brittle fracture. Most fractures are caused by excessive bending, which stresses both aspects of the bone approximately equally. The fact that the side under tensile stress generally fails first indicates that compact bone is better able to resist compression. However, spongy bone is commonly crushed and impacted by compression.
FIG. 1.16
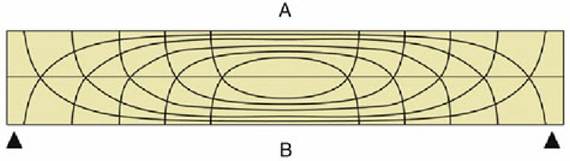
Pattern of compressive (A) and tensile (B) stress lines in a beam supported at both ends. The
greatest stresses (indicated by closeness of lines) occur in the middle of the beam toward the surfaces.
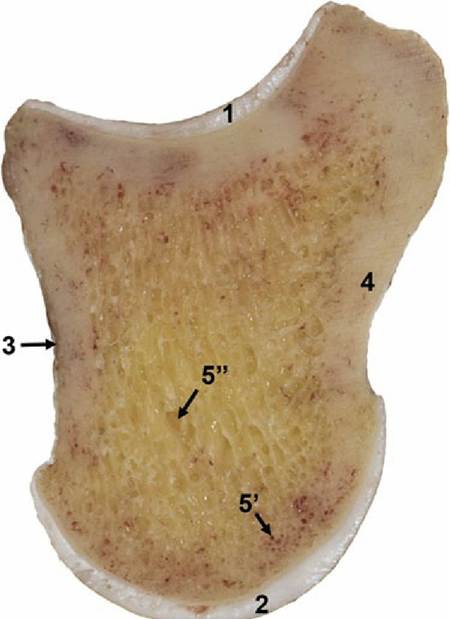
FIG. 1.17 Median section of the unfixed fresh specimen of the second phalanx of equine foot showing articular cartilage at the proximal extremity (1) and distal extremity (2), thin layer of periosteum (3). Notice the change in thickness of the articular cartilages. Cortical compact bone (4) forms the periphery of the bone, and central spongy bone (5) is filled with red (5,) and yellow (5”) bone marrow.
Some Specialized Varieties of Bones
Bones are often found within tendons (rarely within ligaments), where they change direction over prominences that would expose them to excessive pressure and friction. These bones, known as sesamoid bones, form regular synovial joints with the major bones with which they are in contact. In addition to preventing tendon wear, a sesamoid bone displaces the tendon farther from the axis of the adjacent joint and increases the leverage exerted by the muscle. The best known example is the patella (kneecap) in the principal extender of the stifle joint (see Figs. 2.63 and 17.3). In the dog, smaller sesamoids also develop in muscles behind the stifle, in the tendons passing behind the metacarpophalangeal joints (at the bases of the digits), and in the extensor tendons within the digits (see Fig. 1.9). These and other lesser sesamoids may be wrongly identified as chip fractures in radiographs. In large animals, one or more additional sesamoids form dorsal to the deep flexor tendon shortly before its insertion on the distal phalanx (or phalanges). In the dog the reaction is limited to the development of a nubbin of cartilage in each branch of the tendon.
The major sesamoids develop in the embryo before movement is possible, and their origin must therefore be genetically determined. They reform after extirpation only if movement is allowed to indicate their development in reaction to an appropriate stimulus in the lifetime of the animal.
Splanchnic bones develop in soft organs, remote from the rest of the skeleton. The most familiar, indeed the only significant, examples in veterinary anatomy are the os penis (and the female equivalent, os clitoridis) of the dog and cat and the ossa cordis found in the heart, especially in the hearts of ruminants.
Certain bones contain air spaces. In mammals, these pneumatic bones are confined to the skull and contain the paranasal sinuses, which communicate with the nasal cavities. The sinuses develop principally after birth, when outgrowths of the nasal mucosa invade certain skull bones and replace the diploe, the spongy bone between the outer and inner layers ("tables") of compacta. The separation of the tables can be very considerable and can lead to a remarkable postnatal remodeling of the skull, best exhibited by cattle and pigs. The postcranial skeleton of birds develops an extensive system of air-filled cavities in communication with the respiratory organs.