Development
The anatomy of the brain is most easily understood by reference to its development. As such, a general account is provided here; additional details are mentioned as appropriate during descriptions of regional neuroanatomy.
The nervous system makes a very early appearance, becoming evident at the embryonic disk stage as an elongated thickening (neural plate) of the ectoderm that overlies the notochord and paraxial mesoderm. The lateral parts of the neural plate are soon raised above the surrounding surface by growth of the underlying mesoderm and form bilateral neural folds that slope toward an axial crease, the neural groove. As the process continues, the edges of the folds become increasingly prominent and then bend inward toward each other; eventually they meet and fuse, converting the neural groove into a neural tube (Fig. 8.9). The tube, which is the primordium of the brain and spinal cord, then sinks ventral to, and is separated from, the overlying nonneural ectoderm, which fuses dorsal to the neural tube to produce a continuous ectodermal layer. At the same time, cells at the crest of the neural folds, at the junction with the nonneural ectoderm, separate from the folds to form continuous cords, the neural crests, that run almost the whole length of the tube at its dorsolateral aspects. Neural crest cells that remain adjacent to the neural tube eventually develop into neurons, which populate peripheral ganglia (i.e., peripheral sensory neurons and neurons of the visceral motor system), and those neural crest cells that migrate away from the neural tube produce neurons of the enteric nervous system, the medullary parts of the adrenal glands, glia, skin melanocytes, and a variety of craniofacial connective tissues.
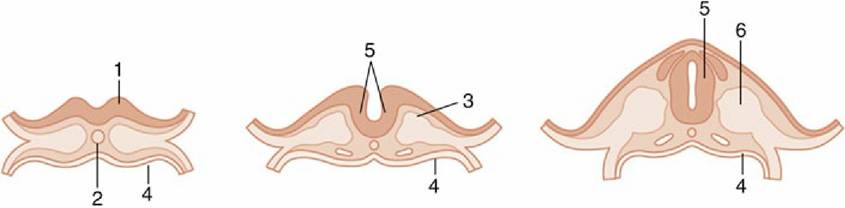
FIG. 8.9 Left to right, Three stages in the closure of the neural plate.
1, Neural plate; 2, notochord; 3, paraxial mesoderm; 4, endoderm; 5, neural tube; 6, somite.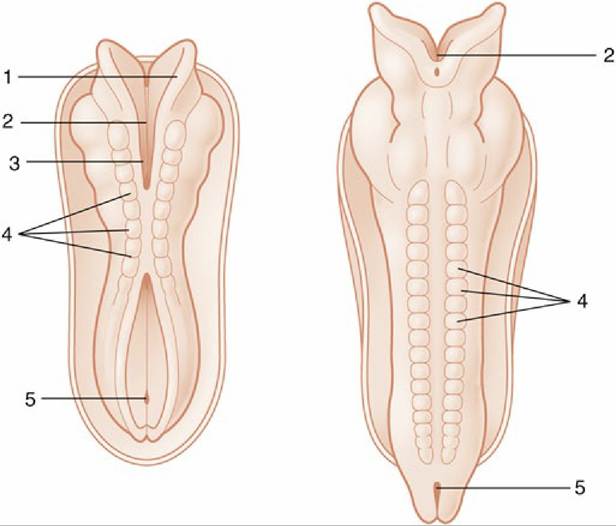
FIG. 8.10 Dorsal views of developing embryos. Two stages in the formation and fusion of the neural folds are illustrated. 1, Neural fold; 2, neural groove; 3, rostral neuropore; 4, somites; 5, caudal neuropore.
Closure of the neural tube initially occurs in the presumptive occipital region, which will become the junction of the spinal cord and brain, but fusion soon spreads rostrally and caudally until only two small openings (neuropores; Fig. 8.10/3 and 5) remain to provide communication at the surface of the embryo between the lumen of the tube and the amniotic cavity. These openings do not persist long: the rostral neuropore closes first, and the caudal one remains open for another day or two while the tube continues to lengthen at its caudal extremity by extension and subsequent infolding of the neural plate. The abnormal persistence of these openings produces relatively common defects of the brain and spinal cord in which nerve tissue may be exposed on the surface of the body. Failure at the rostral extremity leads to malformation of the forebrain and midbrain with accompanying anomalies of the skull; it is known as anencephaly, and although the term implies complete failure of brain development, it can show considerable variation in severity. Most forms are incompatible with life after birth. Failure at the caudal extremity is more common and is known as spina bifida. It is associated with defective closure of the vertebral arches. Children and young animals with this malformation may live after birth, though with severe functional disturbance; affected animals are not usually permitted to survive.
The part of the neural tube that forms the brain is wider from the outset and shows localized expansions even before the tube is completely closed. These define three primary brain vesicles: prosencephalon (forebrain), mesencephalon (midbrain), and rhombencephalon (hindbrain).
The remaining, more uniform part of the tube becomes the spinal cord. The differentiation of the wall of the neural tube is initially similar along the length of the tube but becomes greatly modified later in the part that becomes the brain, increasingly so toward its rostral extremity. The spinal cord develops more uniformly, and its differentiation is considered first.A transverse section of the tube at its formation reveals three concentric layers in its structure (Fig. 8.11A). These are unequally developed around the circumference, which is divisible into thick lateral parts connected by thinner dorsal and ventral regions known as the roof plate and the floor plate. The innermost layer (Fig. 8.11A/1) bounding the lumen of the tube is a continuous layer of neuroepithelial cells—these cells are similar to those that remain as the ependyma lining the adult derivatives of the lumen, the central canal, and the ventricular system of the adult spinal cord and brain. During early development, these neuroepithelial cells proliferate rapidly, and although some daughter cells remain in place adjacent to the lumen, most cells migrate outward into the middle (mantle) layer of the wall of the neural tube (Fig. 8.11A/3). These migrating cells are neural stem cells, precursors of neurons and glia. The mantle layer itself becomes the gray matter of the brain and spinal cord, containing neuronal cell bodies and innumerable neuronal processes. Some of these processes—namely, the axons of developing neurons—grow laterally in the tube, extending outward to form the outer (marginal) layer of the neural tube. The marginal layer (Fig. 8.11A/4) eventually becomes the white matter of the spinal cord, in which fibers descend or ascend along the length of the spinal cord for various distances.
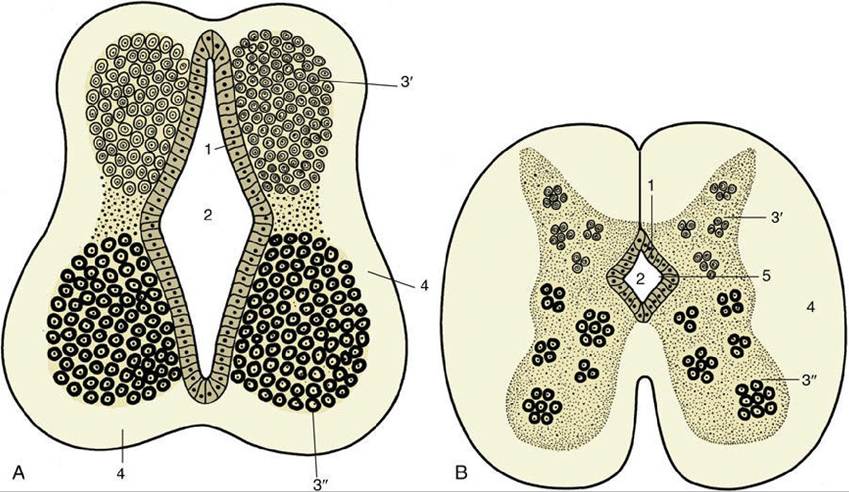
FIG. 8.11 (A) Differentiation of the neural tube. 1, Neuroepithelial (ependymal) layer; 2, central canal; 3' and 3", mantle layer (3', dorsal column, alar lamina; 3", ventral column, basal lamina); 4, marginal layer.
(B) Further differentiation of the neural tube (spinal cord). 1, Neuroepithelial layer; 2, central canal; 3', dorsal column of mantle layer; 3", ventral column of mantle layer; 4, marginal layer; 5, longitudinal limiting groove, or sulcus limitans.
As the number of cells of the mantle layer expands through cell division and migration, the mantle layer becomes arranged in dorsal and ventral columns that bulge into the lumen of the tube, columns that are separated by a longitudinal limiting groove (Fig. 8.11B/5). The dorsal bulge (alar plate) provides the dorsal horn or dorsal column of the gray matter of the cord; its constituent neurons are those receiving synaptic input from sensory or afferent neurons, among other inputs. The ventral bulge (basal plate) becomes the ventral horn or ventral column of spinal gray matter, which is the location of cell bodies of motor or efferent neurons; both dorsal and ventral horns also contain many interneurons. Neurons with somatic functions segregate from those with visceral functions, and four groups of neurons are then arranged in dorsoventral sequence: somatic afferent, visceral afferent, visceral efferent, and somatic efferent (Fig. 8.12). The roof and floor plates provide passages for nerve fibers that pass from one side of the cord to the other, known as commissures.
Further growth of the alar and basal plates causes the lateral parts of the tube wall to expand outward in all directions, submerging the roof and floor plates and creating the dorsal median sulcus and the ventral median fissure that divide the adult cord into its right and left halves. A serial segmentation along the rostrocaudal length of the spinal cord is created by the appearance of the dorsal and ventral roots associated with the spinal nerves. The dorsal roots develop as newly forming axons of developing sensory neurons, differentiating from local condensations of neural crest cells located just lateral to the cord. The axon processes of these sensory neurons extend medially into the cord to reach and penetrate the outer marginal layer.
Branches of these axons can extend over several segments before entering the mantle layer to terminate on neurons in the developing dorsal columns; some branches do not synapse within the spinal cord but turn cranially and extend within the marginal layer to reach the developing brain (see Fig. 8.7). The ventral roots are formed by axons of developing motor neurons within the ventral columns. These axons extend laterally through the marginal layer to emerge on the surface of the cord. The appearance of the dorsal roots on the dorsolateral surface of the spinal cord and of the ventral roots on the ventrolateral surface of the cord divides the white matter into dorsal, lateral, and ventral funiculi(Fig. 8.13/7-9).
Although the cellular development of the nervous system is not described, two points must be made. In most parts of the brain and spinal cord, the full complement of neurons is established shortly after, if not before, birth. However, contrary to former beliefs, in some regions there is a significant, more protracted postnatal recruitment of neural stem cells in areas such as the cerebellum and hippocampus that continues into later life. There is strong evidence that neural stem cells persist, albeit in small numbers, in the adult stages and can be recruited to produce new neurons under certain conditions. The second point relates to the process of myelination of axons within the central nervous system. Different tracts within the brain and cord acquire adequate insulation (essential to their function) at different stages of development, including well after birth. There are important species differences in this process.
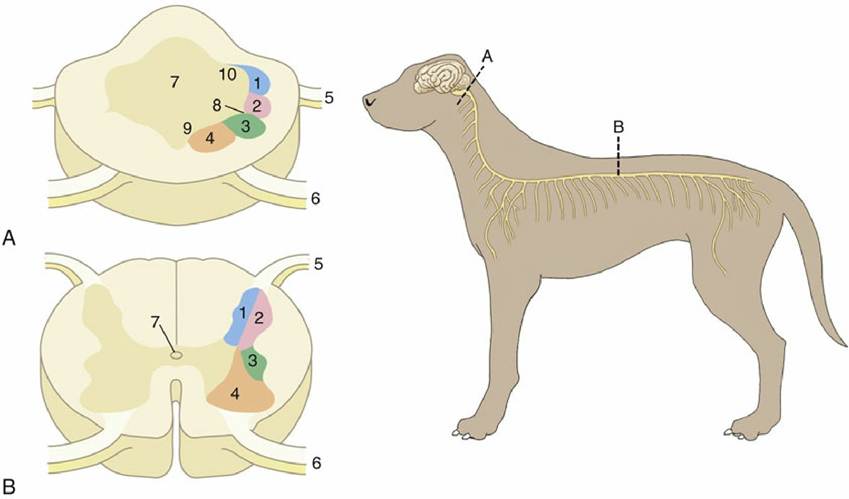
FIG. 8.12 Organization of the gray substance of the (A) medulla oblongata and (B) spinal cord. 1,
Somatic afferent column; 2, visceral afferent column; 3, visceral efferent column; 4, somatic efferent column (lower motor neurons); 5, dorsal root; 6, ventral root; 7, central canal or fourth ventricle; 8, sulcus limitans; 9, basal lamina; 10, alar lamina.
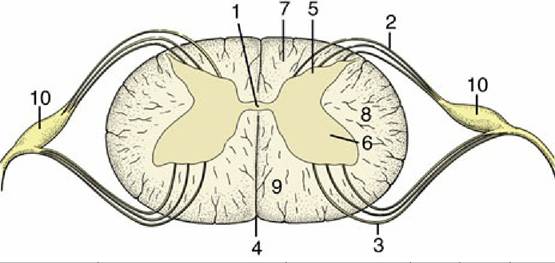
FIG. 8.13 Transverse section of spinal cord showing the subdivision of the white substance by the dorsal and ventral roots of the spinal nerves. 1, Central canal; 2, fibers of dorsal root; 3, fibers of ventral root; 4, ventral median fissure; 5, dorsal horn; 6, ventral horn; 7, dorsal funiculus; 8, lateral funiculus; 9, ventral funiculus; 10, dorsal root ganglion.
During the early development of the brain, three primary brain vesicles, the prosencephalon (forebrain), mesencephalon (midbrain), and rhombencephalon (hindbrain), are evident before closure of the neural tube. At this time, the prosencephalon or forebrain has already extended the evaginations that become the optic cups and eventually form the retina of the eyes. The brain grows more rapidly than the nonneural tissues that surround and enclose it in the embryo, and these constraints enforce a remodeling of its form such that three flexures appear. The most caudal flexure bends the brain ventrally at its junction with the cord. A second flexure at midbrain level occurs almost simultaneously and is sufficiently pronounced to bring the ventral surfaces of the forebrains and hindbrains close together; this relationship is later reversed by the third, or dorsal, flexure, which folds the hindbrain dorsally on itself (Fig. 8.14). The formation of the major divisions of the brain is completed by the appearance of paired lateral evaginations from the alar region of the prosencephalon. These outgrowths, the future cerebral hemispheres, constitute the telencephalon; the unpaired median portion of the prosencephalon, hereafter known as the diencephalon, differentiates into the adult thalamus and related structures. The telencephalon expands in all directions but chiefly in a curve that extends dorsally and caudally to overlap the diencephalon (see Fig. 8.32).
Directly caudal to the developing telencephalon and diencephalon, the midbrain, or mesencephalon, remains undivided. The alar region at this level becomes the tectum of the midbrain, and the basal region the tegmentum of the midbrain. The hindbrain develops cranially into the metencephalon and caudally into the myelencephalon. The basal plate of the metencephalon develops into the structures of the pons in the adult, whereas the alar plate undergoes a bilateral expansion dorsally, becoming the cerebellum. The myelencephalon develops into the medulla oblongata of the adult, continuous caudally with the spinal cord.
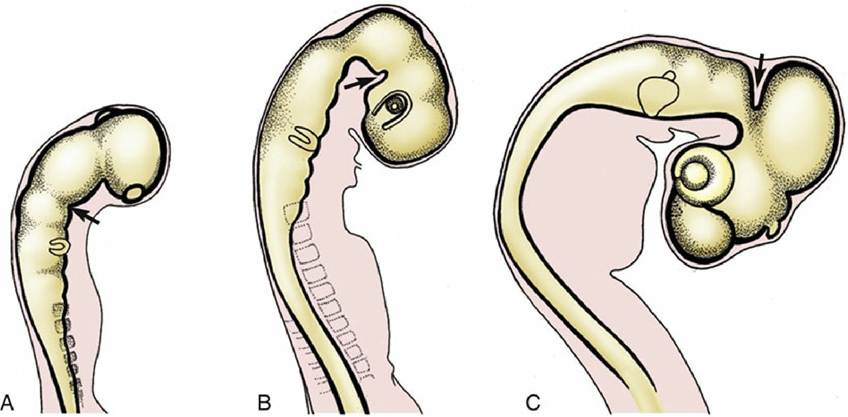
FIG. 8.14 Formation of the (A) caudal ventral, (B) rostral ventral, and (C) dorsal flexures (arrows).
k TABLE 8.1
Derivatives of the Neural Tube
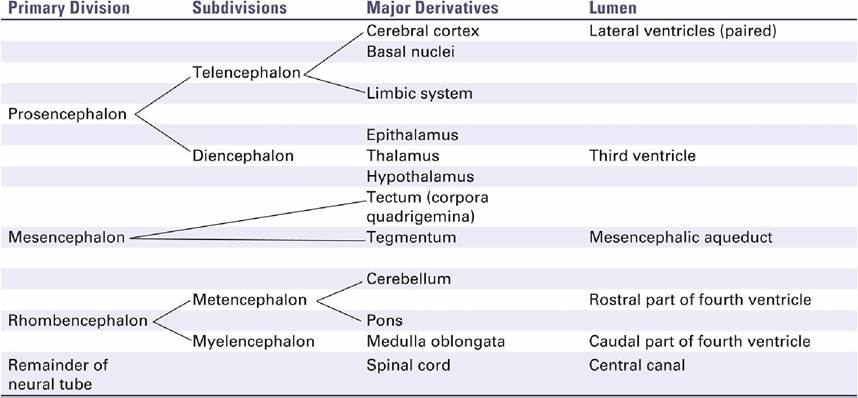
Throughout neural development, the lumen of the original neural tube persists and undergoes modification in shape and size along with the flexures and expansions of the neural tube, eventually becoming the canal and ventricular system of the adult central nervous system. The origin of the major components and ventricles of the brain may be conveniently summarized in tabular form (Table 8.1).