Muscle Contraction and Its Initiation and Cessation Depend on the Binding Specificity and Allosteric Properties of Proteins
There are three types of muscle tissue in vertebrates: skeletal muscle, responsible for the animal’s ability to move; cardiac muscle, a muscle type found only in the heart but structurally similar to skeletal muscle; and smooth muscle, which surrounds hollow organs such as blood vessels, gut, and uterus.
All three produce tensile force by contracting and shortening the length of the muscle. All muscle contraction occurs by the binding and the allosteric properties of two proteins, actin and myosin. Starting and stopping the contraction process depends on two additional proteins in skeletal and cardiac muscle, troponin and tropomyosin. Contraction initiation and cessation in smooth muscle depend on a different system with different proteins and are discussed later in this chapter.Myosin is a large protein whose shape resembles a two- headed golf club. The elongated tail of the myosin molecule corresponds to the shaft of the golf club, and there are two knobs at one end of the tail that, as with golf clubs, are called heads. Myosin tails bind specifically to other myosin tails, forming bipolar aggregates called thick filaments (Figure I-3). Myosin heads specifically bind ATP and another muscle protein, actin. Actin binds to itself to form long, thin filaments, called thin filaments in muscle and called microfilaments or F-actin (filamentous actin) in other cell types. Actin filaments play an important architectural role in all animal cells. Although actin is best understood in muscle cells, all animal cells depend on actin filaments for their shape and for their capacity to migrate in their environment. Actin filaments can be “woven” in various ways to produce different structures, such as ropelike bundles and clothlike networks. These actin bundles and actin networks are used to support the cell in particular shapes, similar to ropes holding up the woven cloth of a tent.
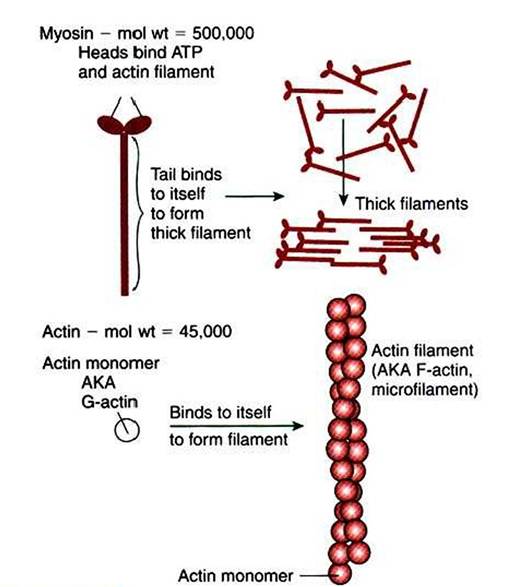
FIGURE 1-3 ■ Assembly of myosin and actin to form filamentous structure.
Myosin tails aggregate with one another to form a thick filament, a substructure of striated muscle. Actin monomers (G-actin) are a single polypeptide chain forming a globular protein that can bind to other actin monomers to form actin filament, also called microfilaments. The actin filament is the basic structure of striated muscle thin filaments; thin filaments also have troponin and tropomyosin as part of their structure.In muscle» the interaction of myosin» ATP, and actin produces contraction and force» as shown in Figure I-4: Step A: ATP binds to a myosin head; in this conformation» myosin has little ability to bind to actin.
Step B: Enzymatic activity associated with the myosin head, an Gdeiiosinetriphosphatase (ATPase), rapidly causes a partial hydrolysis of ATP to adenosine diphosphate (ADP) and inorganic phosphate (Pi), both of which stay bound to the myosin. With ADP and P1 bound, myosin has a slightly different shape that binds avidly to nearby actin filaments.
Step C: When myosin binds to actin, called cross-bridging, the myosin head couples the complete hydrolysis of ATP to a forceful flexing of the myosin head. This allosteric change causes the actin filament to slide past the thick filament. This sliding puts the actin filament under tension, which in turn causes the muscle to contract (shorten) against the load of the muscle (i.e., lifting a weight or pumping out blood). All muscle contraction depends on sliding of actin and myosin filaments. This same allosteric change of myosin also alters myosin-binding properties so that it releases the ADP and P1.
Step D: The binding of a new ATP molecule to the myosin head again causes myosin to change shape; the head unflexes and loses its affinity for actin, releasing the crossbridge, and the cycle can start over. Rigor mortis of dead animals is caused by a lack of new ATP to bind to myosin heads. In the absence of ATP, myosin heads remain in Step C (i.e., bound to actin).
The muscle is stiff because it is completely cross-bridged together.The actomyosin motor uses the binding and allosteric properties of proteins to (I) create structural filaments
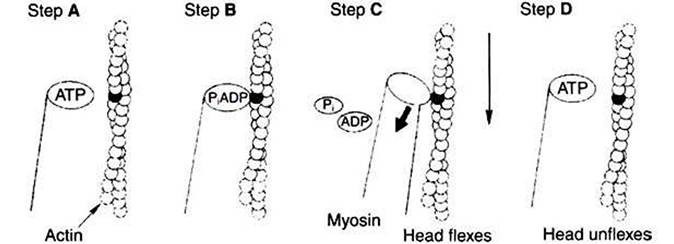
FIGURE 1-4 Power stroke of actomyosin. A,The myosin head has bound to adenosine
triphosphate (ATP). In this conformation, myosin has little affinity to bind to actin. B, ATP is partially hydrolyzed to adenosine diphosphate (ADP) and inorganic phosphate (Pi); the hydrolysis is partial because the products remain bound to the myosin head.The change in what is bound to the myosin (ADP and Pi, not ATP) has the conformation of myosin so that it binds to actin with high affinity. C, Hydrolysis is complete; myosin releases ADP and Pl.This change in what is bound at the myosin head causes an allosteric change in the head; it flexes. Because the myosin head is still bound to the thin filament, the flexion causes the thin filament to slide past the thick filament. D, New ATP molecule binds to the myosin head; as for step At myosin had little affinity for actin in this state, and the head releases from the thin filament and unflexes.
capable of withstanding and transmitting mechanical force, (2) catalyze the hydrolysis of ATP, and (3) couple the “downhill” ATP hydrolysis to the “uphill” contraction to produce force. For just the one protein, myosin, there are a number of examples of the characteristic sequence described earlier: specific binding → protein shape change → change in protein-binding properties and protein function → this change makes a difference.
This system of contractile proteins requires some control so that, for example, the heart beats rhythmically and skeletal muscle contraction is coordinated. At the organismal level, skeletal and cardiac muscle contraction is primarily under control by electrical stimulation from nerves or other electrically active cells (see Chapter 6).
The transmission of electrical excitation to the actomyosin system is called excitation- contraction coupling. Excitation-contraction coupling in all types of muscle depends on changes in intracellular calcium ion (Ca2f) concentration. In skeletal and cardiac muscle, but not smooth muscle, two additional thin-filament proteins, troponin and tropomyosin, are required for this coupling. (Excitationcontraction coupling for smooth muscle is discussed later in this chapter.) In striated muscles, troponin binds to tropomyosin and to Ca2+. Tropomyosin is a long, thin protein that binds in the groove of the actin filament in such a way that its positions, high in the groove or snuggled down deep in the groove, allow or prevent the myosin head access to the thin filament (Figure 1 -5). Excitation-contraction coupling of striated muscle works as follows:Step A: Electrical excitation of a striated muscle cell causes an increase in the intracellular concentration of Ca2+.
Step B: The additional Ca2' binds to troponin, causing an allosteric change in troponin.
Step C: Because Ca2f is bound to troponin, which in turn is bound to tropomyosin, the Ca2+-induced change in troponin conformation is transmitted to the tropomyosin molecule. When troponin binds Ca2f, tropomyosin changes its binding to actin in such a way that it exposes the actin site for myosin cross-bridging. (Tropomyosin snuggles down deeper in its actin groove, revealing actin to the myosin head.) As long as troponin binds Ca2f, the muscle contracts by the actomyosin cycle outlined earlier.
Step D: When the Ca2‘ concentration drops to normal, however, troponin no longer binds Ca2+. This causes tropomyosin to move up in the thin filament groove so that it again blocks the myosin-binding sites on actin. Myosin heads can no longer cross-bridge, and muscle contraction stops.
As with the actomyosin force generation itself, its regulation also shows many examples of the specific binding function.
The specific binding of Ca2+ to troponin is a purely informational use of protein binding and shape change; that is, troponin has no catalytic, transport, or structural function, but transmits the “on” signal to the next protein. The binding of tropomyosin to actin serves not only a regulatory role but also a structural role; the actin filament is stabilized by tropomyosin, making it less likely to disassemble into actin subunits. The change in the binding geometry of tropomyosin that directly regulates myosin access to actin is a good example of the importance of allosteric change and the following sequence: specific binding (troponin to tropomyosin) → protein (tropomyosin) shape change → change in proteinbinding properties (tropomyosin to actin) —> a difference in the position of tropomyosin, which in turn regulates the actomyosin motor.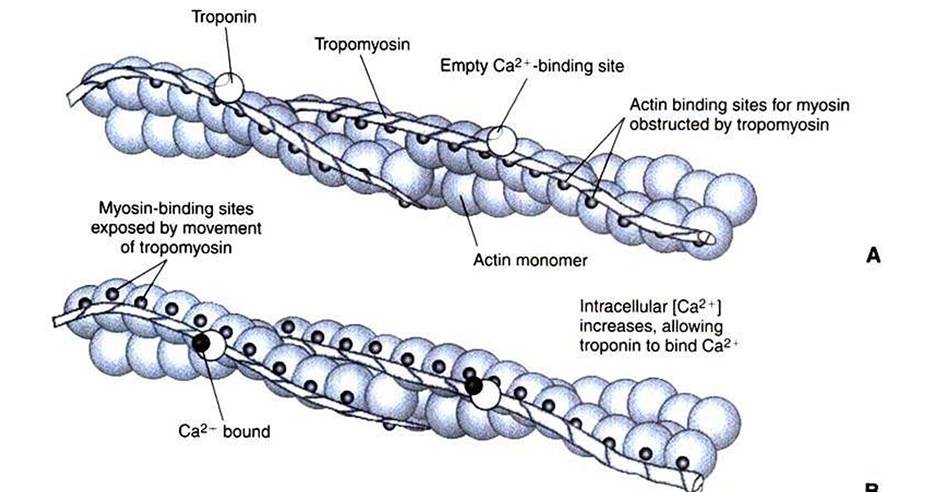
FIGURE 1-5 Regulation of the actomyosin ATPase and striated muscle contraction by Ca2*. A, In the absence of high concentrations of Ca2*, tropomyosin sits in the groove of the actin filament to obstruct the binding sites on actin for myosin. B, In the presence of higher Ca24 concentrations, the ion binds to troponin, causing an allosteric change in the interaction of troponin with tropomyosin. This allosteric change in turn changes the interaction of tropomyosin with the actin filament to expose the myosin-binding sites on actin.