Stimulus-Response Function of the Nervous System
Having established these fundamental points, we may now return to consider the stimulusresponse apparatus in more detail (see Fig. 8.2). Each neuron in the nervous system is part of one or multiple circuits that have specific behavioral functions.
Although not all behavior can be seen as a direct response to a sensory stimulus, we can consider that behavior is produced ultimately as a result of processing of sensory stimuli. This processing can occur either immediately when stimuli are detected—that is, as simple reflex movement—or as a result of longer term processing of previously received and integrated sensory stimuli — stored as memory, for example—which can contribute to self-initiated, more complex behavior. Nevertheless, it is valuable to consider reflexive behavior first, because the underlying neural circuitry contains many of the basic elements of more complex behavior and is itself useful in a clinical context.The monosynaptic reflex arc is often described as the most simple reflex, in that only two neurons, and the synaptic connection between them, are involved in the reflex circuit. This is actually an uncommon arrangement, in that all other known reflexes involve multiple neurons. The monosynaptic reflex, generally termed the stretch or myotatic reflex, is associated with most muscles, the most familiar example of which is the patellar or knee-jerk reflex (Fig. 8.5). This reflex is the rapid, brief contraction of the quadriceps femoris muscle elicited by an appropriate tap on the patellar ligament, the functional continuation of the quadriceps femoris (1). The tap stretches the muscle and stimulates receptors termed muscle spindles within the muscle belly (2), which detect changes in length; activation of the muscle spindle generates action potentials in the sensory afferent neurons (3), innervating the spindle, and these impulses are transmitted along sensory axons within the femoral nerve to reach the spinal cord.
Within the spinal cord gray matter, the sensory neurons form excitatory synapses onto those motoneurons that innervate the quadriceps muscle (4). The axons of these motoneurons in turn exit the spinal cord and travel in efferent direction within the femoral nerve, (5) to innervate the muscle fibers of the quadriceps muscle (6). Motoneuron excitation results in muscle fiber excitation and contraction to produce the brief extension of the stifle joint. This reflex is useful clinically and is elicited when the animal is not weight bearing, in order to produce detectable joint extension. Its essential function for the animal, however, is to maintain extensor muscle activation when the animal is weight-bearing, such that the limbs remain extended against gravity (see Fig. 8.48).In all reflexes other than the stretch reflex, neurons are interposed in the circuitry between the afferent and efferent neurons (Fig. 8.6/3). These neurons are conventionally known as interneurons, although several synonyms exist. Furthermore, all reflexes, including the stretch reflex, involve complex collateral circuitry in which additional neurons not directly interposed between the afferent and efferent neurons are stimulated or inhibited. This arrangement is enabled by the presence of collateral branches of sensory afferent axons within the central nervous system such that the initial sensory signal is divergently distributed within this system and is used by multiple neural circuits (see Figs. 8.7 and 8.48). The purpose of such divergence includes more refined control over the reflex movements and notification of higher centers as to the state of the afferent activity, among other functions.
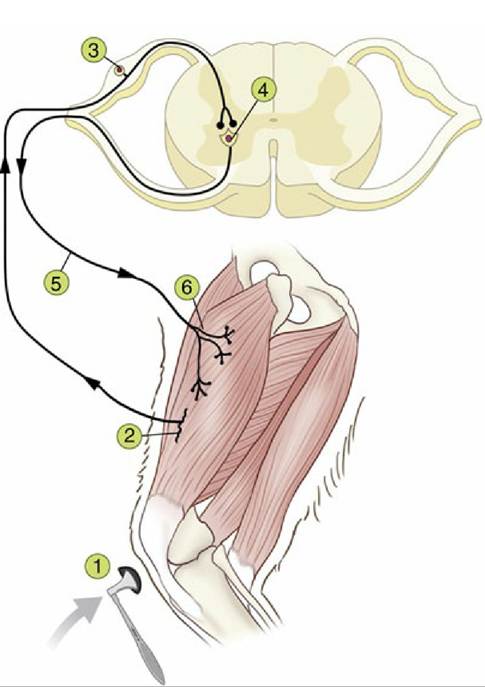
FIG. 8.5 The monosynaptic patellar reflex. The stretch on the tendon (1) stimulates muscle spindles (2), generating action potentials that travel via the afferent neuron (3) to the spinal cord. The impulse is then transmitted to the efferent neuron (4), whose axon (5) innervates the quadriceps muscle (6).
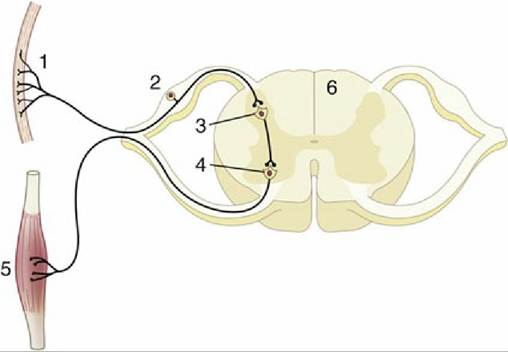
FIG. 8.6 Schematic representation of a reflex chain in which an interneuron is interposed. 1, Skin receptor; 2, afferent neuron; 3, synapse on interneuron; 4, synapse at efferent neuron; 5, muscle; 6, spinal cord.
A good example of such a refined and integrated response is provided by the response to a noxious stimulus applied to the limb of a standing animal. The noxious stimulus is detected by sensory neurons specialized for this modality, termed nociceptors. The reflex response, termed the withdrawal or flexor reflex, is the flexion of all joints of the stimulated limb, such that the limb is withdrawn from the offending stimulus. This action occurs through the coordinated contraction of many flexor muscles across several joints of the limb. The signal from nociceptor activation is distributed within the spinal cord by collateral branching of nociceptor afferent axons, each branch of which synapses upon an individual interneuron, which in turn transmits this excitation to multiple flexor motoneurons (Fig. 8.7/2). Furthermore, the smooth flexion of the limb joints is facilitated by the relaxation of antagonistic extensor muscles that would normally be active in the standing animal. This process necessitates, again through divergence of the same afferent signal, excitation of a different set of interneurons that are capable of inhibiting extensor motoneurons of the same limb. The pathways involved in producing this coordinated activation and inhibition of appropriate muscles extend through several segments of the cord to reach and excite or inhibit the efferent neurons that supply the flexor and extensor muscles of the stimulated limb (Fig. 8.7/3). At the same time, the animal has to adjust to the removal of support from one of its limbs by redistributing its weight over the other limbs; the pathways necessary for this wider adjustment extend through considerable stretches of the spinal cord, some of which cross to the contralateral side (Fig.
8.7).In addition to divergence and integration of afferent information within the spinal cord to coordinate these movements, some information travels cranially to reach brain regions specialized for equilibrium, so that the animal can maintain its balance while standing on three legs. To this complexity of the reflex response is added the inevitable relay of initial nociceptor activation to those regions of the brain involved in decision making, such that the animal can assess the situation and consider whether a more general response, such as flight from or retaliation against the aggressor, would be appropriate. This considered response involves integrative circuitry of various degrees of complexity, extending through the cord and brain, and drawing on those higher centers that are concerned with memory and judgment. Thus, the relationship between reflexive movements and more complex cognitively driven behavior is a continuum, a function of the integration and processing of multiple afferent inputs by increasingly sophisticated neural circuitry.
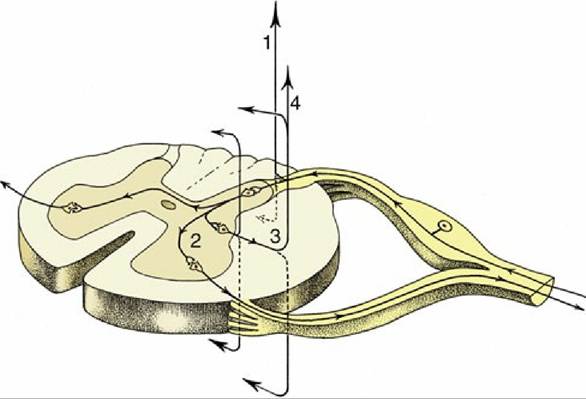
FIG. 8.7 The course of fibers within the spinal cord. Some afferent fibers in the dorsal funiculus travel directly toward the brain (1); others end on interneurons in the dorsal horn. From here impulses can be transmitted directly to efferent neurons (2) or to other interneurons that transmit impulses caudally or cranially within the spinal cord (3), some extending to the brain (4).