THE HINDBRAIN
The hindbrain (rhombencephalon) comprises the medulla oblongata, pons, and cerebellum. These parts differentiate from the caudal brain vesicle shortly after closure of the neural tube.
Attenuation of the roofplate weakens the structure and causes the vesicle to flatten as the pontine flexure develops. The flattening splays the side walls outward so that the luminal surfaces come to face dorsomedially; the alar plates are now lying lateral to the basal plates (see Figure 8-24). The part caudal to the flexure (myelencephalon) becomes the medulla oblongata of adult anatomy. The rostral part develops to become metencephalon, externally marked by the pons and cerebellum. The parts of the roofplate caudal and rostral to the cerebellum remain thin and constitute the medullary vela that complete the enclosure of the lumen, now known as the fourth ventricle (see Figure 8-24).The Medulla Oblongata and Pons
The medulla oblongata and pons form successive portions of the brainstem. The pons corresponds in extent to the large transverse bar that encloses the ventral and lateral aspects and continues into the cerebellum as the middle cerebellar peduncles (see Figure 8-23/9). Despite the clear external distinction, continuity of the internal organization makes the division of pons from medulla a rather artificial concept.
Although the medulla oblongata continues the spinal cord directly, it widens toward its rostral end as the result of the developmental flattening. Its ventral surface is marked by a median fissure continuous with that of the cord and flanked by longitudinal ridges, the pyramids (Figure 8-19/77). Many of the constituent fibers of the pyramids decussate at the transition of spinal cord and medulla, forming interlacing bundles within the fissure. A lesser transverse ridge, the trapezoid body, crosses the ventral surface of the medulla oblongata directly caudal to the larger pontine bar.
The other noteworthy features on this surface are the superficial origins of many of the cranial nerves. The trigeminal nerve (V) appears at the lateral aspect of the transverse pontine bar; the abducent nerve (VI) emerges caudal to this and more medially, through the trapezoid body lateral to the pyramid; the facial (VII) and vestibulocochlear (VIII) nerves appear to continue the trapezoid body laterally; the glossopharyngeal (IX), vagus (X), and accessory (XI) nerves arise from the lateral aspect of the medulla oblongata in close succession; and the hypoglossal nerve (XII) takes a more ventral origin in line with that of the abducent nerve and the ventral roots of the spinal nerves (Figure 8-19 and Figure 8-21).It may be helpful to study a median section (Figure 8-22) of the brain before examining the dorsal aspect of the medulla oblongata and pons. This section shows that the fourth ventricle is brought close to the upper surface of the brainstem by a dorsal inclination of the central canal within the short caudal part of the medulla. The ventricle is covered by a tented roof formed by the cerebellum and the rostral and caudal medullary vela (Figure 8-22/15,15'), which extend from the cerebellum to the closed caudal part of the medulla oblongata and to the midbrain, respectively. Exposure of the dorsal surface of the medulla and pons requires the removal of the cerebellum by transection of its peduncles, which is an operation that almost inevitably destroys the fragile vela (Figure 8-23).
The fourth ventricle is diamond-shaped and is aptly named the rhomboidal fossa; it has its widest part at the pontinomedullary junction. The margins of the fossa are provided by the three pairs of cerebellar peduncles. The floor is rather irregular and is marked by a median sulcus and paired lateral (limiting) sulci. The most
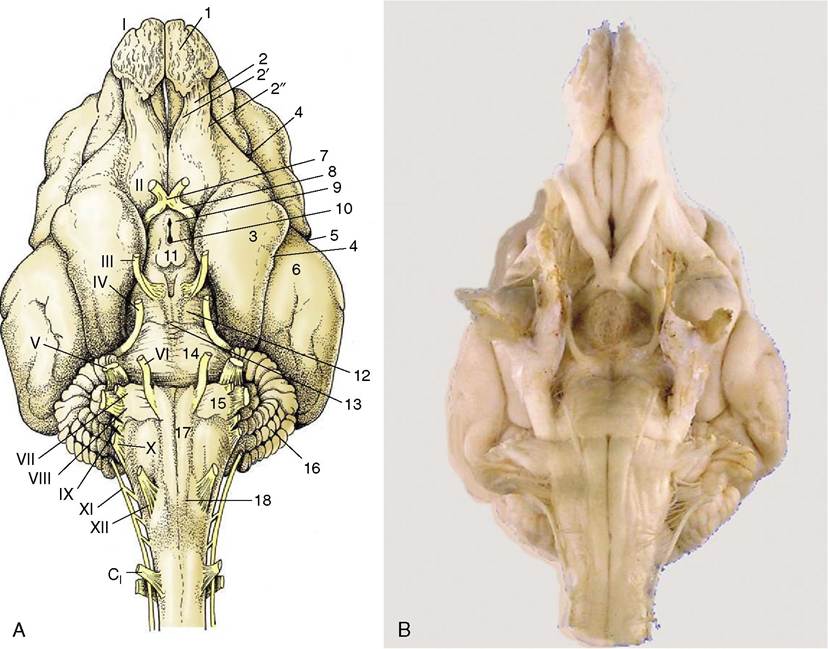
Figure 8-19 A, Ventral view of the canine brain.
1, Olfactory bulb; 2, olfactory tract; 2', medial olfactory tract; 2", lateral olfactory tract; 3, piriform lobe; 4, rhinal sulcus; 5, sylvian sulcus; 6, ectosylvian gyrus; 7, optic chiasm; 8, optic tract; 9, tuber cinereum; 10, infundibulum (the hypophysis has been detached and the third ventricle is opened); 11, mamillary body; 12, crus cerebri; 13, interpeduncular fossa; 14, pons; 15, trapezoid body; 16, cerebellar hemisphere; 17, pyramidal tract; 18, crossing of pyramidal tracts. I-XII designate the appropriate cranial nerves. B, The real specimen of the dog.rostral part of the rostral velum, a part that commonly survives removal of the cerebellum, shows the superficial origins of the trochlear nerves (IV), the only nerves to emerge from the dorsal aspect of the brain.
In the lateral floor of the fourth ventricle and close to the midline the locus coeruleus shines through. Its blue color is due to the presence of neuromelanin granules formed by the polymerization of norepinephrine.
The dorsal surface of the medulla oblongata flanking the caudal part of the fourth ventricle presents inconspicuous eminences, the gracile and cuneate nuclei (Figure 8—20/5,7), at the termination of the like-named fasciculi within the dorsal funiculus of the spinal cord.
The principal features of the internal anatomy of the medulla oblongata and pons are as follows: the nuclei of the cranial nerves, the olivary and pontine nuclei, and the reticular formation and certain ascending and descending fiber tracts that connect the spinal cord with higher levels within the brain. The various categories of structure are described seriatim but without excessive attention to establishing their topographical relationships.
The Nuclei of the Cranial Nerves
The nuclei of the cranial nerves represent the continuation of the four functional components, somatic afferent, visceral afferent, visceral efferent, and somatic efferent, that compose the gray matter of the spinal cord (see Figure 8-12), supplemented by two additional components, special somatic afferent and special visceral afferent, that appear in the medulla oblongata in connection with the innervation of structures of the head that have no counterparts in the trunk or limbs (Figure 8-24).
The first four components are massed together within the gray matter of the cord but separate into parallel columns within the medulla (see Figure 8-12). In part, this is the consequence of the flattening and the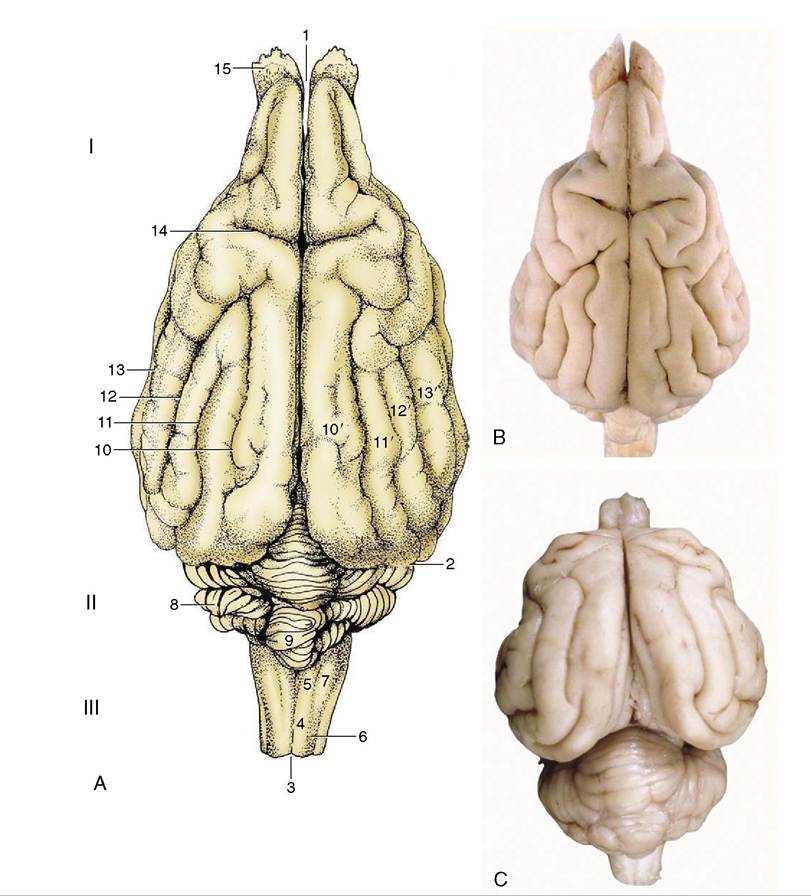
Figure 8-20 A, Dorsal view of the canine brain. I, Cerebral hemispheres; II, cerebellum; III, medulla oblongata. 1, Longitudinal fissure; 2, transverse fissure; 3, dorsal median sulcus; 4, tractus gracilis; 5, nucleus gracilis; 6, tractus cuneatus; 7, nucleus cuneatus; 8, cerebellar hemisphere; 9, cerebellar vermis; 10, marginal sulcus; 10', marginal gyrus; 11, ectomarginal sulcus; 11', ectomarginal gyrus; 12, suprasylvian sulcus; 12', suprasylvian gyrus; 13, ectosylvian sulcus; 13', ectosylvian gyrus; 14, cruciate sulcus; 15, olfactory bulb. B, The real specimen of the dog. C, The real specimen of the cat.
widening of the medulla and dorsal shift in the position of its lumen.
These components now exhibit a lateromedial rather than dorsoventral sequence with a lateral somatic afferent column and a medial somatic efferent column. Certain of the columns also fragment into discrete parts (nuclei), while at some levels the relationships are further adjusted to allow the intrusion of the additional components. The consequences of all this are that those cranial nerves that contain more than one functional component arise from more than one nucleus and that certain nuclei give rise to similar components of more than one nerve. The general arrangement of the six components is illustrated in Figure 8-25 in a schematic fashion sufficient for most needs.
The somatic efferent column serves muscles that have originated from somites and branchiomeres of the head. Its medial part is fragmented into a long hypoglossal nucleus and a smaller abducent nucleus within the floor of the fourth ventricle (and trochlear and oculomo-
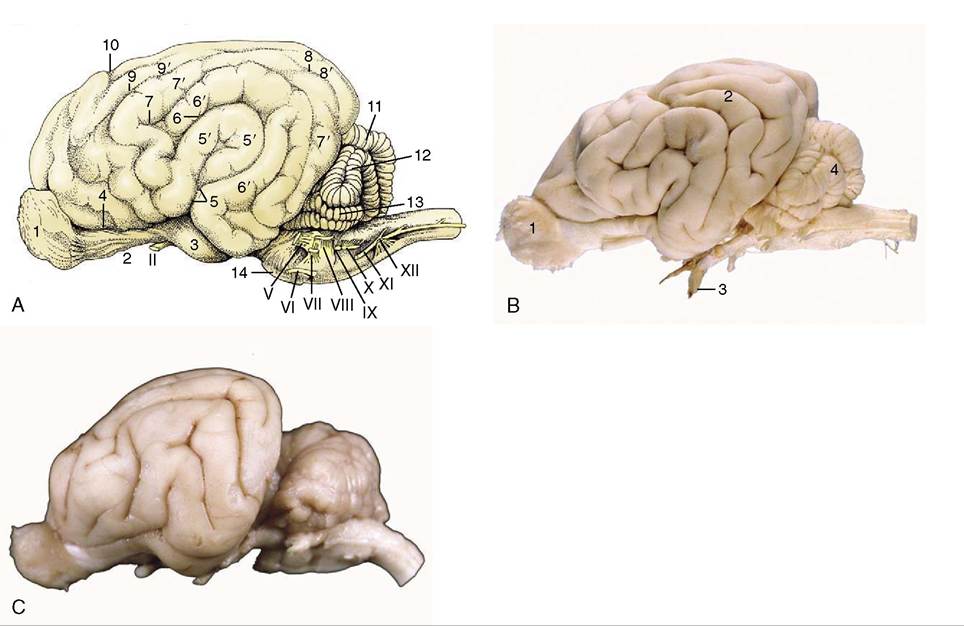
Figure 8-21 A, Lateral view of the canine brain.
1, Olfactory bulb; 2, olfactory tract; 3, piriform lobe; 4, rhinal sulcus; 5, sylvian sulcus; 5', sylvian gyrus; 6, ectosylvian sulcus; 6', ectosylvian gyrus; 7, suprasylvian sulcus; 7, suprasylvian gyrus; 8, ectomarginal sulcus; 8', ectomarginal gyrus; 9, coronal sulcus; 9', coronal gyrus; 10, cruciate sulcus; 11, cerebellar vermis; 12, cerebellar hemisphere; 13, paraflocculus; 14, pons. B, Lateral view of the canine brain. 1, Olfactory bulb; 2, ectosylvian gyrus; 3, optic nerve; 4, cerebellar hemisphere. C, Lateral view of the feline brain.tor nuclei within the tegmentum of the midbrain). The fibers from the oculomotor, abducent, and hypoglossal nuclei take the expected courses to emerge on the ventral aspect of the brain, close to the midline and in line with each other and the ventral roots of the spinal nerves (Figure 8-19). Those that compose the trochlear nerve emerge from the dorsal aspect of the brain after decussation within the rostral medullary velum (Figure 8-23//P); it is an aberrant course for which there is no satisfactory explanation.
The lateral (branchiomeric) portion of the somatic efferent column (see Figure 8-25) supplies the striated masticatory, mimetic, laryngeal, and pharyngeal muscles through the trigeminal, facial, glossopharyngeal, vagus, and accessory nerves. This portion is divided into the motor nuclei of the trigeminal and facial nerves (Figure 8-25/16,17) and the nucleus ambiguus (Figure 8-25/14) shared by the glossopharyngeal and vagus nerves. The fibers emerge from the ventrolateral surface of the brainstem but do not always take the most direct internal course to do so.
The visceral efferent column supplies the autonomic (parasympathetic) motor component of certain cranial nerves. It is the lateral of the efferent columns (Figure 8-24/4) and is divided into the parasympathetic nucleus of the vagus (Figure 8-25/13), the caudal salivatory nucleus of the glossopharyngeal, and the rostral salivatory nucleus of the facial nerve (Figure 8-25/15) (and the parasympathetic nucleus of the oculomotor nerve [Figure 8-25/15] in the midbrain).
The distribution of the vagal fibers of this category is to the cervical, thoracic, and abdominal (but not pelvic) viscera, and the distribution of those within the glossopharyngeal and facial nerves is to glands of the head (and the distribution of those within the oculomotor nerve is to intrinsic muscles of the eyeball).The visceral afferent column (Figure 8-24/2,3) is in fact double and is shared by visceral and special visceral afferent neurons. It forms a single very long nucleus (of the solitary tract [Figure 8-25/10]) that is subdivided in relation to the associated facial, glossopharyngeal, and vagus nerves. Many neurons are concerned with visceral
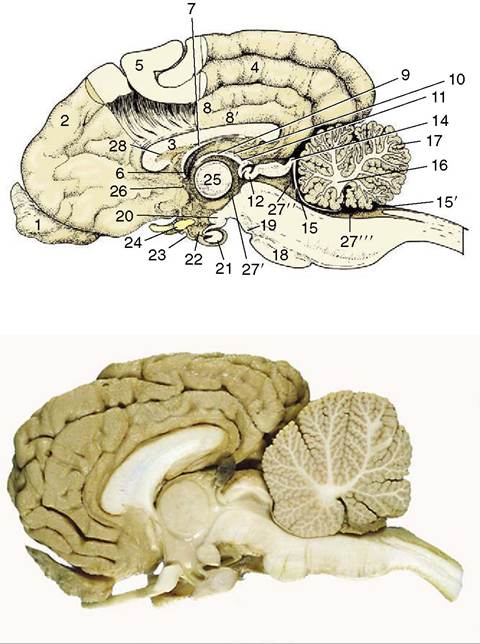
Figure 8-22 Median section of the canine brain. Part of the medial wall of the hemisphere has been removed. 1, Olfactory bulb; 2, hemisphere; 3, corpus callosum; 4, splenial sulcus; 5, cerebral cortex; 6, interventricular foramen; 7, fornix; 8, cingulate gyrus; 8', supracallosal gyrus; 9, thalamus; 10, epithalamus; 11, epiphysis; 12, posterior commissure; 13, 14, commissures of rostral and caudal colliculi; 15, rostral medullary velum; 15', caudal medullary velum; 16, corpus medullare; 17, cerebellar cortex; 18, pons; 19, crus cerebri; 20, mamillary body; 21, hypophysis; 22, infundibulum; 23, tuber cinereum; 24, optic chiasm; 25, interthalamic adhesion; 26, anterior commissure; 27’, third ventricle; 27", mesencephalic aqueduct; 27", fourth ventricle; 28, septum telencephali (pellucidum).
sensation in the caudal part of the mouth and the cervical, thoracic, and abdominal viscera; the special component, which is concerned with taste, is spread between all three named nerves.
The somatic afferent column (Figure 8—24/1) extends from the cervical part of the spinal cord through the medulla and pons into the mesencephalon. It is broken into several nuclei. One, the mesencephalic nucleus of the trigeminal nerve (Figure 8-25/7), is concerned with proprioception; it presents a unique feature, the inclusion of the primary afferent neuron cell bodies within the central nervous system (the one exception to an otherwise inviolable rule that the cell bodies of primary afferent neurons are located within peripheral ganglia). The two exteroceptive nuclei (Figure 8-25/7) are the nucleus of the descending (spinal) tract of the trigeminal nerve, which extends from the level of the nerve’s entrance into the cervical part of the spinal cord, and the principal sensory nucleus of the trigeminal nerve within the pons.
The special somatic afferent column is associated with the optic and vestibulocochlear nerves and therefore with the special somatic senses of vision (II), balance (vestibular division of VIII), and hearing (cochlear division of VIII) (Figure 8-25/6,8,9). The afferent pathways of these important senses are considered elsewhere; our present purpose is to locate the relevant nuclei within the brainstem. The four closely related vestibular nuclei are spread through part of the medulla oblongata and pons, medial to the caudal cerebellar peduncle. The two (dorsal and ventral) cochlear nuclei are located within the most rostral part of the medulla oblongata close to the entry of the eighth nerve.
The fiber composition of the nerves are summarized conveniently within Table 8-2.
Other Internal Features
The olivary nuclear complex occupies a position in the caudal part of the medulla oblongata, dorsolateral to the pyramidal tract, where it sometimes raises a gentle surface swelling (Figure 8-26/10). It is composed of several parts and varies considerably in form among species, generally taking the form of a nuclear lamina folded onto itself to form a bag. It is an important feature of the motor feedback regulatory mechanism (pp. 302-303). Several other nuclei within the pons (Figure 8-27) are also concerned with motor control (p. 301).
The reticular formation is a diffuse system of nuclei and fiber tracts (Figures 8-26/8 and 8-28/13) that extends from the spinal cord to the forebrain and occupies a large part of the core of the medulla oblongata and pons. It is discussed on p. 298.
The principal fiber tracts that pass through this part of the brainstem also receive attention later. The large descending tract that produces the pyramid externally (Figure 8-26/11) and the ascending tract known as the medial lemniscus (Figure 8-28/9) are prominent in transverse sections. The medial lemniscus is formed of fibers that issue from the gracile and cuneate nuclei, run ventrally (as the deep [internal] arcuate fibers), and cross the midline in the ventral part of the caudal medulla before turning rostrally as a large medial lemniscal bundle. This area also includes fibers of the trigeminothalamic and cervicothalamic tracts, which emanate
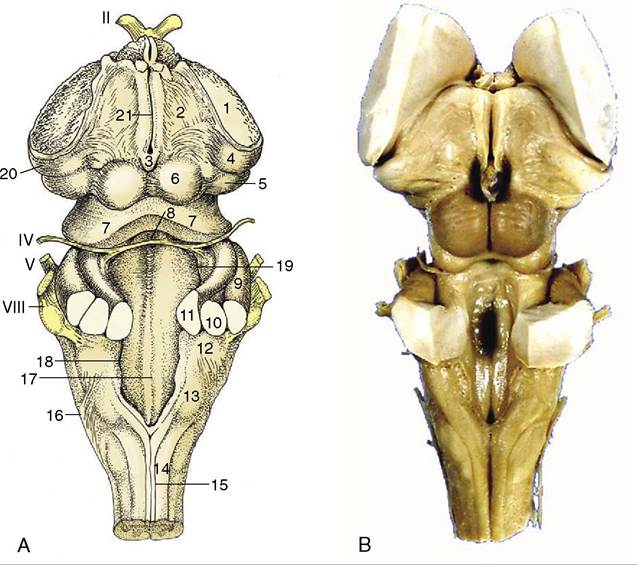
Figure 8-23 A, Dorsal view of the canine brainstem with the cerebellum removed and the fourth ventricle opened. 1, Cut fibers of internal capsule; 2, dorsal part of thalamus; 3, epiphysis; 4, lateral geniculate body; 5, medial geniculate body; 6, rostral colliculus; 7, caudal colliculus; 8, decussating fibers of trochlear nerves in the rostral velum; 9, middle cerebellar peduncle; 10, caudal cerebellar peduncle; 11, rostral cerebellar peduncle; 12, dorsal cochlear nucleus; 13, cuneate tubercle; 14, fasciculus cuneatus; 15, fasciculus gracilis; 16, superficial arcuate fibers; 17; median sulcus; 18, medial eminence; 19, sulcus limitans; 20, optic tract; 21, margin of roof of third ventricle. B, Dorsal view of equine brainstem.
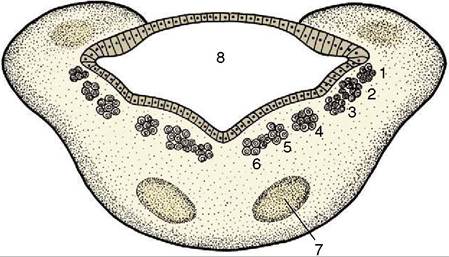
Figure 8-24 Schematic transverse section of the metencephalon. The special somatic afferent nuclei are not shown. 1, Somatic afferent column; 2, visceral afferent column; 3, special visceral afferent column; 4, visceral efferent column; 5, 6, somatic efferent column; 7, nuclei of pons; 8, fourth ventricle.
from the principal sensory nucleus of the trigeminal nerve and the lateral cervical nucleus, respectively. Other conspicuous fiber aggregations compose the three cerebellar peduncles whose composition, origin, and destination are given later.
The Cerebellum
The cerebellum is a roughly globular, much-fissured mass that is located above the pons and medulla oblongata and is connected to the brainstem by three peduncles on each side (Figure 8-23/9,10,11). It is separated from the cerebral hemispheres by the transverse fissure occupied by the membranous tentorium cerebelli (p. 308) when the brain is in situ.
The cerebellum consists of large lateral hemispheres and a narrow median ridge named the vermis from its fancied resemblance to an earthworm. A division of
greater functional and phylogenetic significance is created by a series of transverse fissures. The deepest divide a small caudal flocculonodular lobe from the larger mass, which is itself divided into caudal and rostral lobes (Figure 8-21). Smaller fissures divide the
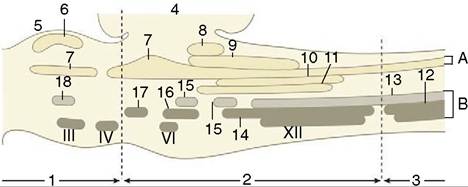
Figure 8-25 Schematic representation of the brainstem showing the nuclei in an adult mammal. Roman numerals are used for nuclei of some cranial nerves. A, afferent nuclei; B, efferent nuclei. 1, Mesencephalon; 2, rhombencephalon; 3, spinal cord; 4, cerebellum; 5, tectum mesencephali; 6, rostral colliculus (SSA); 7, trigeminal nuclei (SA); 8, cochlear nuclei (SSA); 9, vestibular nuclei (SSA); 10, solitary nucleus of VII, IX, X (VA); 11, gustatory nuclei of VII, IX (SVA); 12, motor nucleus of XI (SE); 13, motor nucleus of X (VE); 14, nucleus ambiguus of IX, X (SE); 15, salivatory nuclei of VII, IX (VE); 16, motor nucleus of VII (SE); 17, motor nucleus of V (SE); 18, parasympathetic nucleus of III (VE). SSA, special somatic afferent; SA, somatic afferent; VA, visceral afferent; SVA, special visceral afferent; SE, somatic efferent; VE, visceral efferent.
lobes into lobules and these into yet smaller units known as folia. The caudal lobe is particularly well developed in higher forms and especially so in primates. The lobules are individually named, but neither their names nor their exact forms are important.
The arrangement of the gray and white substance sharply contrasts that found in the spinal cord and medulla oblongata. In the cerebellum the bulk of the gray substance is arranged as an external cortex that encloses the white substance or “medulla” (Figure 8-22). The medulla arises from the peduncles and radiates through the various lobes, lobules, and folia, forming a branching structure with some resemblance to a tree. Because of this appearance and because of an ancient belief that it is the seat of the soul, it is sometimes known as the arbor vitae—the tree of life. Some additional gray substance forms a series of nuclei embedded within the medulla; the most important of these are the fastigial nuclei (Figure 8-27/13) close to the midline, the lateral cerebellar (dentate) nucleus (Figure 8-27/15) laterally, and the nuclei interpositi (Figure 8-27/14).
The cerebellum is attached to the brainstem by the three cerebellar peduncles on each side and by the caudal and rostral medullary vela (see Figure 8-23). The caudal peduncle (Figure 8-23/10) connects with the medulla oblongata and is largely composed of afferent fibers, of which some run from origins within the spinal cord and others run from the vestibular nuclei, the olivary nucleus,
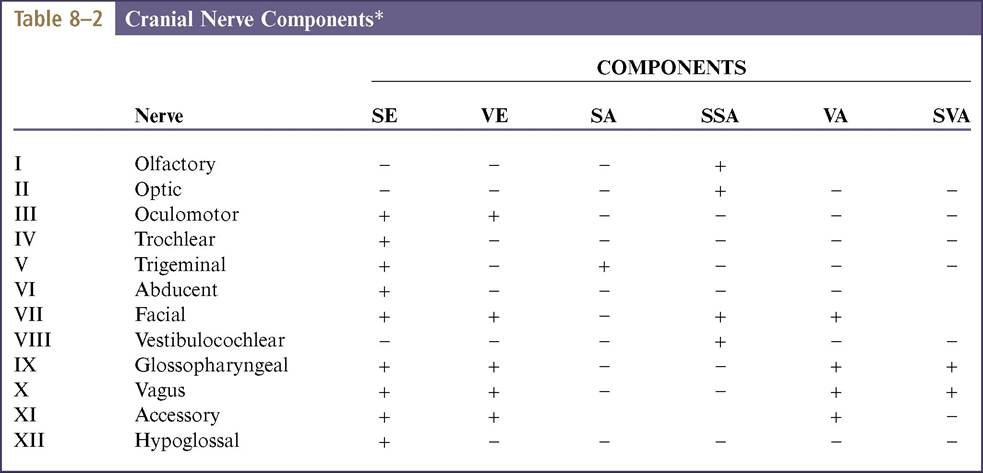
*Certain points are controversial: notably the nerve trunks followed by fibers conveying proprioceptive information from various muscles of the head, and the precise distribution of the medullary component of the accessory nerve.
SE, Somatic efferent; VE, visceral efferent; SA, somatic afferent; SSA, special somatic afferent; VA, visceral afferent; SVA, special visceral afferent.
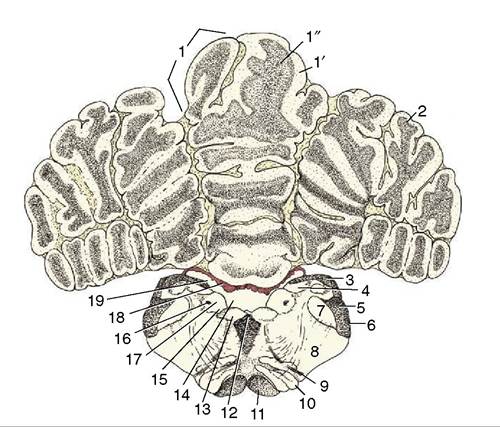
Figure 8-26 Transverse section of the canine brain at the level of the hypoglossal nerve (XII). 1, Cerebellar vermis; 1', cortex; 1", medulla; 2, cerebellar hemisphere; 3, fasciculi gracilis and cuneatus; 4, gracile and cuneate nuclei; 5, caudal cerebellar peduncle; 6, spinal tract of the trigeminal nerve; 7, nucleus of the spinal tract of the trigeminal nerve; 8, reticular formation; 9, root of hypoglossal nerve; 10, caudal olivary nucleus; 11, pyramidal tract; 12, medial longitudinal tract; 13, motor nucleus of XII; 14, sulcus limitans; 15, motor nucleus of X; 16, solitary tract (special visceral afferents of VII, IX, and X); 17, solitary nucleus; 18, choroid plexus; 19, fourth ventricle.
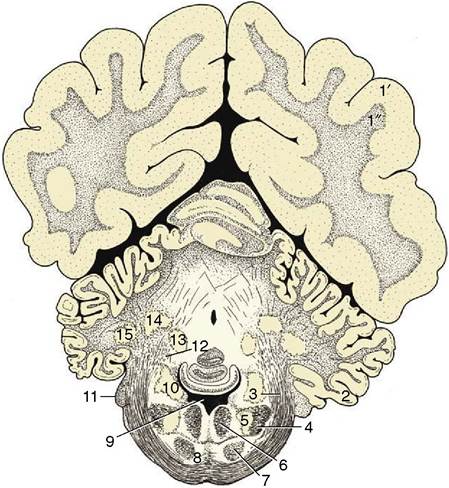
Figure 8-27 Transverse section of the canine brain at the level of the middle cerebellar peduncle. 1', 1", cerebral hemisphere; 1', neocortex; 1", fibers; 2, paraflocculus lateralis; 3, middle cerebellar peduncle; 4, spinal tract of the trigeminal nerve; 5, nucleus of the spinal tract of the trigeminal nerve; 6, medial longitudinal fasciculus; 7, pyramidal tract; 8, pontine nuclei; 9, fourth ventricle; 10, nuclei of the vestibulocochlear nerve (VIII); 11, root of VIII; 12, rostral cerebellar peduncle; 13, fastigial nucleus; 14, nucleus interpositus; 15, lateral cerebellar nucleus.
and the reticular formation. The middle peduncle (brachium pontis; Figure 8—23/9) is also composed of afferent fibers; these arise from pontine nuclei. The rostral peduncle (brachium conjunctivum; Figure 8—23/11) is attached to the midbrain; it is largely composed of efferent fibers dispatched toward the red nucleus, reticular formation, and thalamus but also includes a considerable afferent component that continues the ventral spinocerebellar tract. The three peduncles are closely compressed together at their attachments to the cerebellum.
The functions of the cerebellum are concerned with the control of balance and the coordination of postural and locomotor activities. Balance is located in the flocculonodular node. The caudal lobe is concerned with the feedback regulation of motor function, and to this end it receives a direct input from pontine and olivary nuclei and an indirect input from the other parts of the cerebellum. The rostral lobe receives an input of proprioceptive information. There is a somatotopic representation of the body in the cerebellar cortex.