When Protein and Energy Availability in the Forestomach Are Well Matched, Rapid Microbial Growth and Efficient Protein Utilization Result
Because a large part of preformed dietary protein is fermented in the rumen, ruminant animals depend, to a large extent, on microbial protein to meet their own protein needs. Microbial protein reaches the abomasum and small intestine when microbes are washed out of the rumen and into the lower tract.
Digestive efficiency is optimized in ruminants when the growth rate of the microbial mass is maximal, resulting in maximal delivery of microbial protein to the host animal. These conditions are best met by rapidly growing populations of microbes. The microbial growth rate depends on the supply of nutrients and the rate at which microbes are washed from the rumen. Here we consider the effect of nutrient supply on microbial growth rate; factors affecting the rale of microbial removal are discussed later.The overall reaction in the rumen may be greatly simplified, for the purposes of this discussion, to Equation 1:
Glucose + Peptide = Microbes + VFA + NH3 + CH4 + CO2 (1) Glucose and peptide represent ruminally available carbohydrate and protein, respectively. In this context, available means available to the microbes for fermentation. Carbohydrate or protein that is not susceptible, or accessible, to microbial attack is classified as unavailable and is not included in Equation 1. Glucose was chosen to represent carbohydrate, and peptide to represent protein, because all carbohydrates must be broken down to simple sugars, and proteins to peptides, before becoming available to bacteria. The term peptide in this equation could be replaced by other forms of nitrogen, but for now, the discussion is confined to peptide as a nitrogen source. Peptide is the only nitrogen-containing substrate on the left in the equation, but there are two nitrogen-containing products on the right: microbes (as protein) and NH3 (ammonia).
Both substrates, glucose and peptide, contain carbon, oxygen, and hydrogen and thus can contribute to the formation of microbial carbon, VFA, CH4, and CO2.Equation 1 always balances, but the distribution of products varies according to the relative concentrations of substrates, as illustrated in Figure 31-5. For microbial cells to be produced, both energy and nitrogen are required. Energy can come from either glucose or peptide, but nitrogen must come from peptide. When glucose and peptide availability are appropriately matched (Figure 31-5, «), energy for cellular growth comes primarily from glucose, with peptides directed toward microbial protein synthesis. Under these conditions, the products of Equation 1 favor microbial cells with little ammonia production. Glucose fermentation with accompanying VFA production must be high to meet the large energy demands necessary to support the rapid growth of the microbial mass. Ammonia production is low because most peptide nitrogen is being incorporated into microbial protein.
When the availability of glucose is high relative to peptide (Figure 31-5, b), there is ample energy but insufficient nitrogen to support adequate protein synthesis, and thus microbial
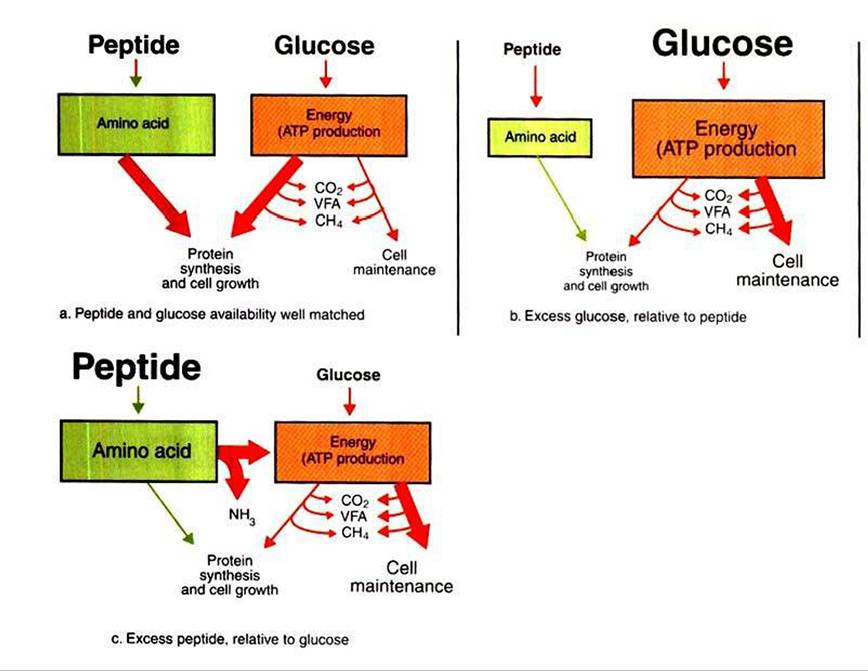
FIGURE 31-5 The efficiency with which dietary energy is used for protein synthesis in the rumen depends on the balance between energy and nitrogen sources.The proportion of energy used for protein synthesis and cell maintenance (as indicated by the size of the arrows) changes in relation to the balance of peptide (nitrogen) and glucose supplies. ATP, Adenosine triphosphate; VFA, volatile fatty acid.
replication is not maximal. In this case, microbial energy utilization becomes inefficient as energy is used for the maintenance of nondividing cells, rather than for the energyrequiring synthetic processes of growing cells.
The maintenance energy needs of the microbes still drive some fermentation of glucose with moderate VFA production, but production of both microbial cells and ammonia is limited because of lack of nitrogen.When peptide availability exceeds glucose availability (Figure 31-5, c), there is ample nitrogen to support growth, but growth is limited because of insufficient energy supplies. These conditions force the microbes to use peptides to meet energy needs instead of for protein synthesis. Microbial growth rate is low, and VFA production is moderate, because fermentation is driven only by the maintenance energy needs of the microbes. Much of the VFA production comes from the carbon portions of the peptides, whereas the amine groups arc shunted to ammonia production; thus the products of Equation 1 favor ammonia.
The relationship between available glucose (carbohydrate) and peptide (or nitrogen) has a tremendous effect on the production of microbial cells and thus a profound impact on the nutrition of the host. This relationship, as illustrated in Figure 31-5, is quantified by expressing microbial growth in terms of grams of microbial dry matter produced per mole of energy-producing substrate used. This value, referred to as microbial yield, is usually designated by a capital Y subscripted ■with the abbreviation of the energy substrate to which it is referenced. A convenient but somewhat theoretical substrate with which to reference microbial cell yield is ATP. Microbial yield is then written as Yλγp = x, where x is the number of grams of microbial dry matter produced per mole of ATP used. The value of Yaγp varies between about 10 and 20 g of microbes per mole of ATP. Nitrogen availability, from either peptide or nonprotein sources, has an important effect on the
Yatp value. When microbial growth is limited by low nitrogen availability, a large portion of available ATP is used for maintenance rather than cell growth; thus the number of cells produced per ATP is small, and the Yλip value is low.