IMMUNITY
Immunity is an animal’s ability to resist harmful organisms and toxins, which is crucial for maintaining health and preventing diseases. There are two primary forms of immunity, namely innate and acquired.
The immune system is a multifaceted structure comprising organs, cells, and proteins that recognise and counteract foreign pathogens such as viruses, bacteria, and toxins. The different parts of the immune system consist of white blood cells, antibodies, and lymphoid organs. The innate immune system reacts quickly and without specificity to a variety of pathogens. While, the adaptive immune system responds specifically to pathogens and retains a memory of the encounter for future protection.5.2.1 Innate Immunity
Innate immunity, or nonspecific immunity, is the body’s first line of defence against pathogens. It is a fundamental immune system component that provides immediate but non-specific defence against infection. The innate immune system has the ability to respond quickly to a wide variety of pathogens, unlike the adaptive immune system, which is more specific and requires time to mount a response.
The innate immune system comprises various physical barriers, cellular components, and chemical factors. The body has natural defences like skin and mucous membranes that protect against the entry of harmful pathogens through respiratory, gastrointestinal, and urogenital systems. These barriers are often supplemented by chemical defences, such as the acidic environment of the stomach and the lysosomal found in bodily fluids like saliva, tears, and reproductive tract pH. Within the respiratory tract, several mechanisms can reduce the antigen load, including the turbulence of air that occurs during inhalation. When microorganisms are carried by this turbulence as well as ciliary action of the respiratory tract, they are pushed onto the walls of the respiratory tract, lined with sticky mucus and bactericidal lysozymes, further reducing the antigen load.
The innate ability of urine and milk to flush out infectious antigens is crucial in preventing infectious conditions. Failure to eliminate urine or milk can result in the development of infections. The gastrointestinal tract has a protective layer of symbiotic bacteria that stops harmful bacteria from sticking to its target cells. This mechanism is similar to the protection provided by the skin. On a cellular level, the innate immune system includes various types of leukocytes, such as macrophages, neutrophils, and natural killer cells, which can engulf and destroy pathogens through phagocytosis. Neutrophils release web-like structures called neutrophil extracellular traps (NETs) during processes like NETosis (Figure 5.2).These NETs are composed of DNA strands associated with histones and are decorated with various proteins, including bactericidal proteins from granules and cytoplasm. When exposed to bacterial lipopolysaccharides (LPS) or proinflammatory cytokines, such as tumour necrosis factor-alpha (TNF-α) and IL-8, neutrophils become activated and initiate the process of NET formation. When tissues are damaged, chemical messengers and chemokines attract neutrophils to the site of infection. This attraction leads to the expression of adhesion molecules on vascular endothelial cells. Neutrophils bind to these cellular adhesion molecules through specific receptors, which trigger them to leave the circulation by crossing capillary walls (diapedesis) and entering the tissues.
In addition to these components, the innate immune system also includes a range of soluble factors, such as the complement system, a group of proteins that can be activated to destroy pathogens, and various antimicrobial peptides and proteins, such as lysozyme, that can directly kill or inhibit the growth of microbes.
Despite its broad range of defences, the innate immune system is not infallible. Pathogens can sometimes evade these defences and establish an infection. However, the innate immune system plays a crucial role in limiting the spread of infection and buying time for the adaptive immune system to mount a more specific and potent response.
5.2.2 Antigens and Their Characteristics
Antigens have the ability to activate lymphocytes, which can cause the body to initiate an immune response. Antibody-binding sites on an antigen are called antigenic determinants or antigenic epitopes. Antigens can be
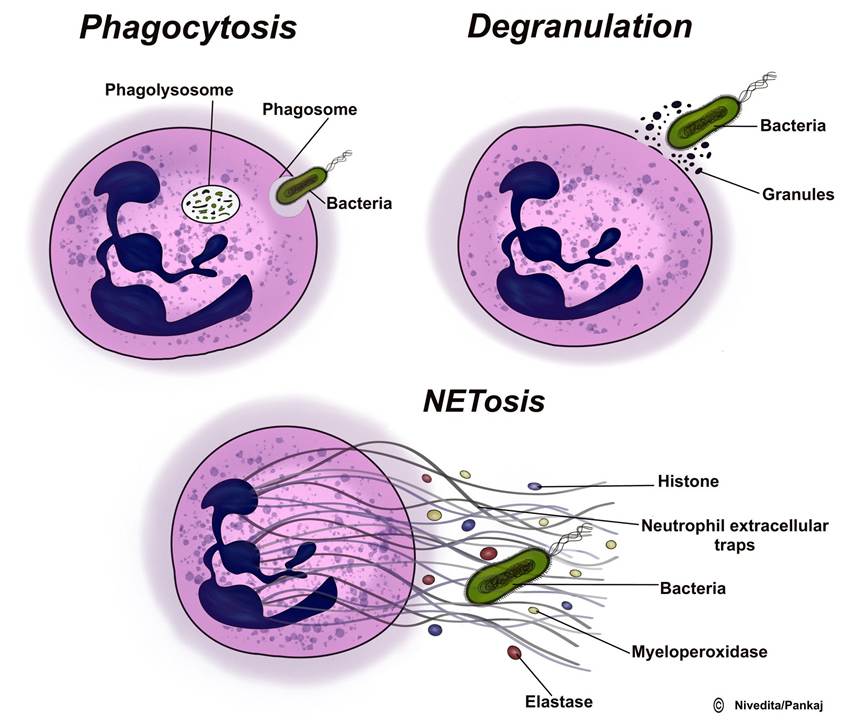
FIGURE 5.2 Different actions of neutrophils to destroy the pathogen
broadly categorised into infectious (microbial) and nonin- fectious antigens. Infectious antigens include components derived from bacteria, viruses, protozoa, and helminths. In contrast, noninfectious antigens encompass a wide range of substances, including those derived from “self,” food, plants, dust, insect and animal venoms, and synthetic and cell surface proteins.
The level of immune response triggered by an antigen is known as antigenicity or immunogenicity. Understanding the features of antigens that can stimulate a strong or weak immune response is important. This knowledge is critical in understanding the body’s ability to fight against invading antigens and developing effective vaccine preparations.
Antigens have different features determining their effectiveness: 1. Biochemical structure: More complex molecules can elicit more robust immunogenic responses than simpler molecules. For example, proteins with small sizes but complex structures have better immunogenicity than simple carbohydrates or lipids.2. If an antigen is bigger, it will get processed more effectively by antigen-presenting cells.3. Flexibility and degradability: If the antigen is less flexible, it can generate varied immunogenic responses. However, strong immunogenic responses can be generated with better stability and more degradability. Foreign antigens are more potent than self or host antigens. Moreover, the immune system’s response to antigens can be affected by genetics, biomolecules that regulate and modulate immune responses, as well as the amount and method of antigen exposure.
5.2.3 Acquired Immunity
Acquired immunity develops over time in response to specific antigens. This type of immunity is characterised by its ability to learn, adapt, and remember, tailoring its response to each specific antigen encountered. Acquired immunity is mediated by specialised immune cells, including B lymphocytes that produce antibodies in a process known as humoral immunity and T lymphocytes responsible for cell-mediated immunity. These immune responses are initiated in the lymphoid tissues of the body. Acquired immunity can be acquired through various means, such as exposure to an infection, vaccination, or transmitting antibodies from another animal. Upon encountering a specific antigen, the immune system creates a memory that enables a faster and more effective response upon future exposure to the same antigen.
5.2.3.1 Lymphocytes and Their Role in Immunity
Despite their similar appearance under a microscope, they are divided into two primary groups: T lymphocytes and B lymphocytes. These cells, originating from multipotent haematopoietic stem cells in the embryo, play distinct roles in the immune system. T lymphocytes, or T cells, are involved in cell-mediated immunity, while B lymphocytes, or B cells, are key players in humoral immunity, producing antibodies. The formation of lymphocytes and the development of cell-mediated and humoral immunity are depicted in Figure 5.3. After preprocessing, both T and B cells migrate to lymphoid tissue throughout the body.
A. T cells and cell mediate immunity: T cells derive their name from the thymus gland, which is the location where they travel to and undergo processing. This preprocessing involves rapid division and the development of diverse reactivity against specific antigens. The thymus gland ensures that T cells do not react against the body’s proteins or antigens, preventing autoimmunity. This preprocessing primarily occurs shortly before and a few months after birth. During their processing, they are transformed into four categories:
1.
Cytotoxic T cells, or CD8+ T cells, are responsible for directly killing infected host cells and tumour cells. Upon activation, cytotoxic T cells induce apoptosis in the target cells, thereby eliminating the source of infection or abnormal cell growth.2. Helper T cells, or CD4+ T cells, are a heterogeneous collection of cells that can transform into different subsets, including Th1, Th2, Th17, and Tfh. Each subset has distinct effector functions and plays a role in coordinating immune responses. For example, Th1 cells activate macrophages and promote inflammation, while Th2 cells are important for activating B cells and promoting antibody production. Th17 cells are associated with the recruitment of neutrophils and the defence against extracellular pathogens. Tfh cells are essential for the development of long-lived antibody responses
3. Suppressor T cells are a specific type of T cell that has an essential function in preserving the immune system’s balance and avoiding autoimmune responses. They suppress other immune cells’ activation and effector functions, preventing excessive immune responses and tissue damage.
4. Memory T cells are a specialised subset of T lymphocytes that provide long-term immunity against previously encountered pathogens. Their ability to recognise and eliminate specific pathogens upon re-exposure is essential for preventing the recurrence of infectious diseases.
B. B lymphocytes undergo maturation in two stages: first, in the liver during mid-foetal life, and then in the bone marrow during late foetal life and after birth. The name “B cells” originates from the bursa of Fabricius, an organ in birds where these cells were first discovered. B cells differ from T cells in two main ways: they actively secrete antibodies that react against antigens and exhibit even greater diversity than T cells, forming millions of types of antibodies with different specific reactivities. The main types of B cells include:
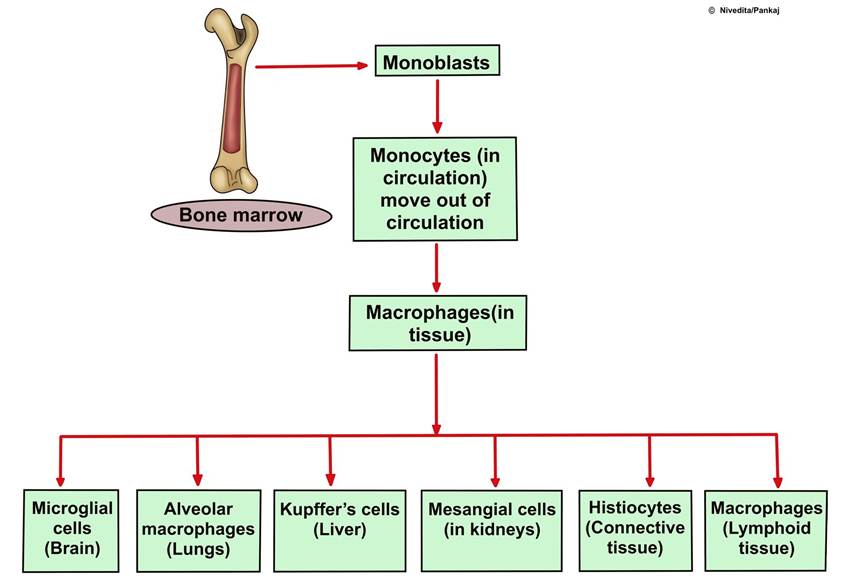
FIGURE 5.3 Formation of lymphocytes and generation adaptive immunity
1.
Plasma Cells: These are the effector cells of B lymphocytes and are responsible for the production and secretion of antibodies. Once activated, B cells differentiate into plasma cells. Plasma cells produce large amounts of antibodies to fight specific antigens.2. Memory B Cells: These long-lived B cells form following an initial immune response to an antigen. The immune system’s long-term defence is aided by memory B cells, which quickly identify and respond to previously encountered antigens upon re-exposure.
3. Naive B Cells: Naive B cells are mature B cells that have not yet encountered their specific antigen. They circulate in the bloodstream and lymphoid organs, waiting to encounter antigens that will activate them.
Humoral immunity: B cells produce antibody molecules, which may be secreted or inserted into the plasma membrane. When an antigen activates a naive or memory B cell, it proliferates and differentiates into an antibody-secreting effector cell, known as a plasmablast or plasma cell. In addition, B cells present antigens and secrete cytokines. B cells can recognise antigens and respond to the antigen in a specific way, expressing specific antibodies on the membrane.
The B-cell population of a specific activated clone significantly increases, and the new B lymphocytes are added to the original lymphocytes of the exact clone. These B cells also travel throughout the body to populate all the lymphoid tissue. However, they do not become active until they encounter another quantity of the same antigen that originally activated them. On subsequent exposure to the same antigen, the memory cells will cause a much more rapid and potent antibody response the second time because there are many more memory cells than the original B lymphocytes of the specific clone. The administration of multiple doses of antigen with intervals of several weeks or months between injections is usually required for effective immunisation because the increased potency and duration of the secondary response make immunisation successful. When naive B lymphocytes encounter their associated antigens, become activated, and undergo clonal expansion, they differentiate into short-lived or long-lived plasma cells that produce large amounts of antibodies. The short-lived plasma cells provide rapid protection but are destroyed after a few days of antibody secretion. However, the long-lived plasma cells reside in tissues such as the bone marrow and gut-associated lymphoid tissue. They can continue producing antibodies for many years, providing lifelong immunity against infectious diseases like measles and smallpox.
Generating these lymphocytes is a complex process regulated by various factors, including chemokines, BCR signalling, and specific antigens. This process ensures a continuous replenishment of the B cell repertoire, essential for maintaining the body’s defence against foreign substances.
5.2.4 Antibodies
Antibodies, also known as immunoglobulins (Igs), are a class of gamma globulins produced by B lymphocytes in response to the antigen. Antibodies recognise and bind to antigens to neutralise and remove them from the body.
There are five general classes of antibodies, each designated by a specific letter: IgA, IgD, IgE, IgG, and IgM. Among these five, 75 % of total antibodies are IgG. IgG is the only antibody class that significantly crosses the animal placenta, providing passive immunity to the foetus. Colostrum produced by cattle after parturition has a high concentration of antibodies, including immunoglobulins IgA, IgG, and IgM, which provide passive immunity to newborns during crucial phases of life. IgE has a role in allergic conditions. All these antibodies play various roles in the immune response, such as localised defence, recognition of antigens, and involvement in allergic reactions.
5.2.4.1 The Antibody's Structure
Comprises two pairs of chains, specifically one pair of heavy or long chains and one pair of light or short chains (Figure 5.4).
Each heavy chain contains about 400 amino acids, while each light chain contains about 200 amino acids. The antibody is divided into two halves, each by disulphide bonds. Each half of the antibody consists of one heavy chain and one light chain, and disulphide bonds join the two chains in each half. Each antibody molecule always has at least two and as many as ten such pairs. The combination of nonco- valent and covalent bonds - disulphide holds the light and heavy chains together.
The antibody contains two regions: the constant region and the variable region.
A. T he constant region, also known as the Fc (Fragment crystallisable) region, is similar in number and placement of amino acids in all antibodies of each type. The constant portion determines other properties of the antibody, such as antibody diffusivity in the tissues, adherence to specific structures in the tissues, attachment to the complement complex, and the ease with which the antibodies pass through membranes.
B. T he variable region is also known as the Fab (Fragment antigen binding) region. The specificity of antibodies results from the unique structural organisation of amino acids in the variable portions of the light and heavy chains. The amino acid organisation has a different steric shape for each antigen specificity, allowing for rapid and tight bonding between the antibody and antigen.
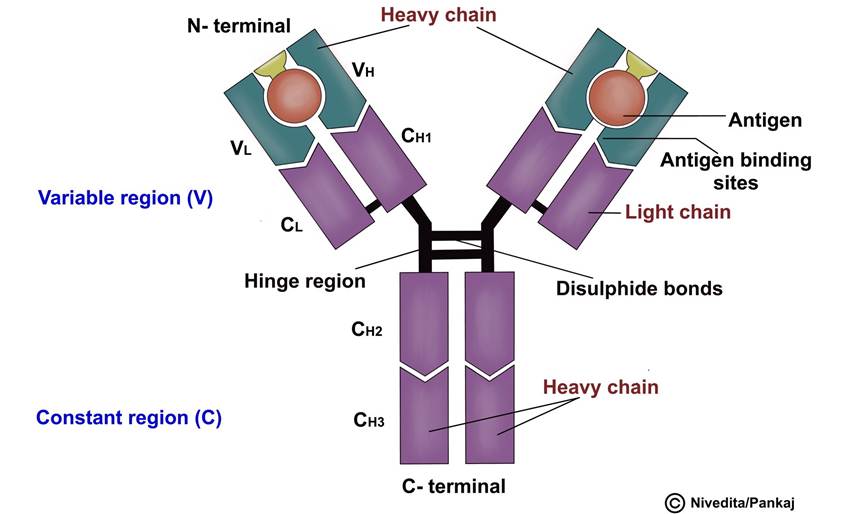
FIGURE 5.4 Structure of an antibody showing two heavy and two light chains
If an antibody is very specific, then there are many sites where it can bond with an antigen. This bonding is firm and is held together by different chemical forces such as hydrophobic bonding, hydrogen bonding, ionic attractions, and van der Waals forces.
5.2.5 Antigen-Presenting Cells
Antigen-presenting cells (APCs) play an important role in detecting, engulfing, and presenting antigens to T cells, triggering an immune response. There are three main types of APCs: macrophages, dendritic cells, and B lymphocytes.
1. Macrophages are large phagocytic cells that digest invading organisms to release antigens. Macrophages are highly versatile and heterogeneous, taking on different names according to their tissue location. Some of the different types of macrophages, based on their tissue location, include Lung-Alveolar macrophages or dust cells, Skin-Langerhans cells, Liver-Kupffer Cells, Central Nervous system-Microglial cells, Kidney Mesangial cells, and connective tissues-Histio- cytes (Figure 5.5). They are present in various lymphoid tissues and are considered the major APCs.
2. Dendritic cells, conversely, are nonphagocytic and classified based on their location, such as in the spleen, lymph nodes, or skin. They are highly efficient in antigen presentation and are considered the most potent APCs.
3. B lymphocytes, traditionally known for their role in antibody production, have also been found to act as APCs, although they are the least efficient among the three types.
The role of APCs is to process and present antigens to T cells, specifically helper T cells. The major histocompatibility complex (MHC) proteins facilitate the interaction between APCs and T cells, which bind peptide fragments of antigen proteins degraded by APCs and transport them to the cell surface. T cells recognise major histocompatibility complexes using their T cell receptors (TCRs). The specific functions of cytotoxic and helper T cells depend on the presentation of antigens by APCs. MHC can be separated into two classes: class I MHC molecules, present on all nucleated cells of the body and responsible for presenting endogenous antigens to cytotoxic T cells, and class II MHC molecules, found on APCs and responsible for presenting exogenous antigens to helper T cells.
The process of antigen presentation by APCs is essential for the specificity and regulation of the adaptive immune response. It allows the immune system to differentiate between self and non-self antigens, thereby mounting an effective response against foreign invaders while preventing autoimmunity. The proper function of APCs and the presentation of antigens are critical for activating and regulating lymphocytes, which are instrumental in the adaptive immune response.
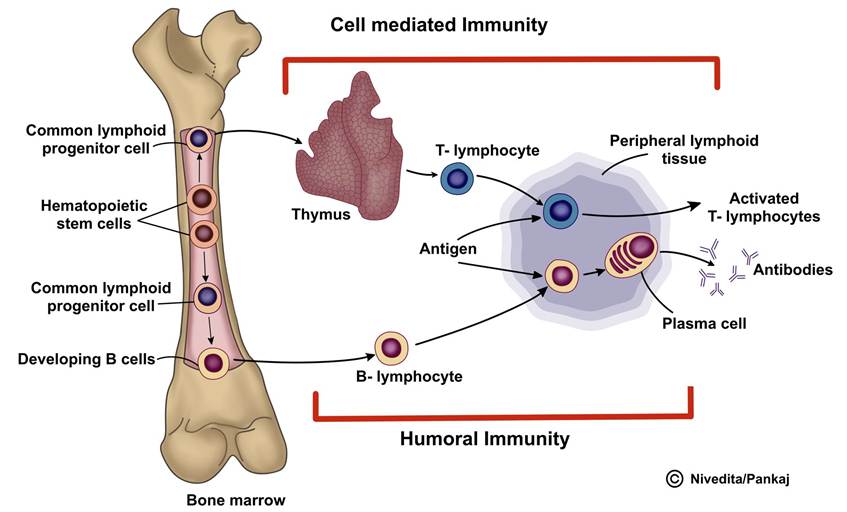
FIGURE 5.5 Diagram shows the formation of monocytes and, upon entry into different tissues, their synonyms
5.2.6 Toll-like Receptors
Toll-like receptors (TLRs) are a group of pattern recognition receptors that are crucial parts of the innate immune system. They recognise specific sequences or molecules in microbes, known as pathogen-associated molecular patterns (PAMPs). TLRs are mainly expressed by innate immune cells, particularly macrophages, and are the primary defence mechanism against invading pathogens. Upon recognition of PAMPs, TLRs trigger innate immunogenicity, leading to the generation of proinflammatory cytokines, chemokines, and antimicrobial peptides, as well as the upregulation of costimulatory molecules.
In mammals, 13 TLRs have been identified, each with a specific role in recognising distinct PAMPs. TLRs 1 to 6, except 3 are located on the cell surface membrane, where they identify a wide variety of microbial products, including bacterial lipoproteins, lipopolysaccharides (LPS), and flagellin. On the other hand, TLRs 3, 7, 8, and 9 are found intracellularly, where they recognise nucleic acids from viruses and bacteria. TLR4 responds to LPS present in tissues and the bloodstream, triggering proinflammatory signalling pathways. TLR4 has been shown to recognise endogenous molecules produced from tissue injury, indicating that TLR4 represents a key receptor on which both infectious and noninfectious stimuli converge to induce TLR4-mediated inflammation.
Toll-like receptors (TLRs) have a role in initiating the immune response to infections and tissue damage. They are also involved in several acute and chronic diseases, having a pivotal role as an amplifier of the inflammatory response.
Their dysregulation can lead to chronic inflammation and contribute to the progression of various diseases.
5.2.7 Mode of Action of Antibodies
Antibodies can protect the animal body by two mechanisms: A. Direct actions and B. Complement pathway, as shown in Figure 5.6.
Direct actions: Because of the bivalent nature of the antibodies and the multiple antigen sites on most invading agents, when antibodies come into contact with these agents, they can deactivate them in various ways. These methods include:
1. Agglutination: Where multiple large particles having antigens on their body (RBCs and bacteria) are clubbed with each other to form a clump.
2. Precipitation: Where the molecular complex of a soluble antigen (e.g., tetanus toxin) and antibody complex is large enough that it becomes insoluble and precipitates.
3. Neutralisation: Where the antibodies cover the toxic sites of the antigenic agent.
4. Lysis: Where some strong antibodies can directly attack cellular agent membranes, causing them to rupture.
These direct actions of antibodies often are not strong enough to play a major role in protecting the body against the invader, but indirect actions provide more potent
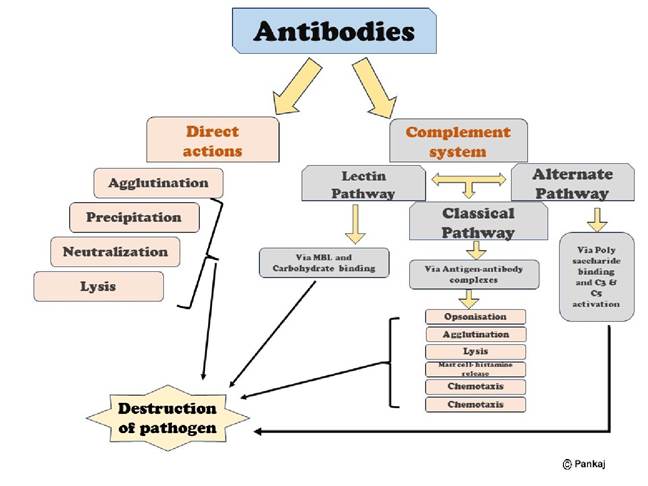
FIGURE 5.6 Mode of action of antibodies: Direct and complement system
protection by activating the complement pathways. A complement system enhances or accelerates various activities during the fight against invading organisms. It is a system of plasma enzymes identified by numbers from C1 to C9. Including the three subunits of C1 (C1q, C1r, C1s), there are 11 enzymes. Usually, these enzymes are inactive and are activated in three ways: a. Classical pathway, b. Lectin pathway, c. Alternate pathway.
a. Classical pathway: The classical pathway of the immune response begins when an antibody interacts with an antigen. This interaction exposes a specific reactive site on the antibody, which then binds with the C1 molecule of the complement system. This triggers a series of sequential reactions, starting with the activation of the proenzyme C1. The activated C1 enzymes then sequentially activate other enzymes in the system, leading to an amplified reaction from a small beginning. This results in multiple end products that help protect the body’s tissues from damage caused by the invading organism or toxin. Key effects include:
i. Opsonisation and phagocytosis, where the complement cascade product C3b significantly boosts phagocytosis by neutrophils and macrophages, engulfing bacteria attached to the antigen-antibody complexes. This process, known as opsonisation, can increase the number of bacteria destroyed several hundred times.
ii. Lysis is another important effect, where the membrane attack complex (also known as the cytolytic complex), a combination of multiple complement factors C5b6789, inserts itself into the cell membrane, creating pores that lead to the osmotic rupture of the cell membranes of bacteria or other invaders.
iii. The complement products also cause agglutination by changing the surfaces of the invading organisms, causing them to stick to each other. They can neutralise some viruses, rendering them nonvirulent.
iv. Chemotaxis is initiated by fragment C5a, leading to a large influx of neutrophils and macrophages into the tissue area near the antigenic agent.
v. When fragments C3a, C4a, and C5a are present, they trigger mast cells and basophils to release various substances, including histamine and heparin, into the nearby fluids. These substances increase regional blood flow and fluid leakage into the tissue, among other local tissue reactions that help inactivate or immobilise the antigenic agent.
vi. Besides the inflammatory effects caused by the activation of mast cells and basophils, other complement products also contribute to local inflammation. These products increase blood flow, further increase capillary leakage of proteins, and cause interstitial fluid proteins to coagulate in the tissue spaces. This prevents the movement of the invading organism through the tissues.
b. The lectin pathway is activated when the serum protein mannose-binding lectin (MBL) binds to the mannose or fructose groups on the surface of bacteria, fungi, or viruses.
c. The alternative pathway is activated by a circulating protein called factor I. This protein binds to polysaccharides present in the cell membrane of invading organisms, leading to the activation of C3 and C5, which ultimately attack the antigenic products of the invading organism.
5.2.8 Cytokines
Cytokines are small proteins that serve as cell signalling molecules, aiding cell-to-cell communication in immune responses and regulating inflammation in the body. They are produced by various cells, including immune cells like macrophages, B lymphocytes, T lymphocytes, mast cells, endothelial cells, fibroblasts, and stromal cells. Cytokines significantly influence the balance between humoral and cell-based immune responses, regulating specific cell populations’ maturation, growth, and responsiveness. They act through cell surface receptors and are involved in autocrine, paracrine, and endocrine signalling. Additionally, they are involved in various processes such as inflammation, pain, host immune responses to infection, trauma, sepsis, cancer, and reproduction.
5.2.8.1 Types of Cytokines
1. Interleukins: Produced by leukocytes, interleukins regulate cell growth, differentiation, and motility, affecting the immune system. Interleukins can be either inflammatory or antiinflammatory, or both types.
2. Interferons: Interferons have antiviral, immunomodulatory, and antiproliferative effects, playing a key role in the immune response against viral infections.
3. Tumour Necrosis Factors (TNF): TNF plays a crucial role in inflammation and can induce fever, apoptotic cell death, and sepsis.
4. Chemokines: These cytokines play a crucial role in the immune response by stimulating the transfer of immune cells to the site of injury or infection.
5. Lymphokines: Lymphokines relay messages between immune cells, contributing to regulating the immune response.
6. Monokines: Monokines help mobilise an immune response and are produced by monocytes and macrophages.
5.2.9 Passive Immunity
Passive immunity refers to giving a person antibodies to a disease rather than producing them through their immune system. This kind of immunity offers instant protection, although it is not long-lasting and may only last for several weeks up to 3 or 4 months. Passive immunity can occur naturally, for example, when antibodies from the mother are passed to the foetus via the placenta or from breast milk to the baby’s gut. It can also be produced artificially, such as when antibodies are given as a medication.
Passive immunity can be further classified into two types: natural and artificial.
1. Natural Passive Immunity: This immunity is obtained from the mother before and after childbirth. During pregnancy, the mother passes immunity to the foetus through IgG antibodies in the placenta. After birth, the antibodies (IgA) are transmitted through milk. The antibodies received from the mother are metabolised soon. Therefore, the passive immunity is short-lived.
2. Artificial Passive Immunity: This type of immunity is developed by injecting previously prepared antibodies using serum from humans or animals. Antibodies are obtained from the persons affected by the disease or from animals, particularly horses, which have been immunised artificially. The antibody (antiserum) serum is administered to animals who have developed the disease (therapeutic). It is also used as a prophylactic measure. This type of immunity helps provide immediate protection against acute infections like tetanus for poisoning by insects, snakes, and venom from other animals.
Unlike active immunity, which can take days or weeks to develop, passively administered antibodies can provide rapid and immediate protection. Passive antibodies are not dependent on the recipient’s immune system, unlike vaccination, which relies on the host’s immune response to create protective immunity. Passive immunisation may thus represent the therapy of choice in highly endemic areas where vaccine coverage is low or in individuals who cannot mount an effective immune response.
5.2.10 Allergy and Hypersensitivity
Immunity, while crucial for protecting the body from harmful pathogens, can sometimes lead to undesirable side effects such as allergies or hypersensitivity reactions. When the immune system overreacts to allergens, it causes allergic reactions. Allergens can be anything from food to physical agents like heat or light. There are several types of hypersensitivity reactions, some of which only occur in animals with a specific predisposition to allergies. One such reaction is anaphylaxis, a severe and widespread allergic reaction that can occur when an allergen enters the bloodstream. This reaction involves basophils in the blood and mast cells in the tissues, which, if sensitised by IgE reagins, can trigger a body-wide allergic response. This response includes the release of histamine, leading to widespread vasodilation and increased capillary permeability, which can result in significant plasma loss from the circulation. If not promptly treated with epinephrine, anaphylaxis can lead to circulatory shock and potentially death.
Another type of hypersensitivity reaction is urticaria, also known as hives, which results from antigens entering specific skin areas and causing localised anaphylactoid reactions. Histamine released in these areas causes vasodilation and increased capillary permeability, leading to localised skin swelling.
Hypersensitivity reactions can be classified into five types: Type I or anaphylactic reactions, Type II or cytotoxic reactions, Type III or antibody-mediated reactions, Type IV or cell-mediated reactions, and Type V or stimulatory/ blocking reactions. Each type of reaction involves different immune system components and varies based on the type of antigen, the immune response generated, and the speed at which the response is produced.
Common allergens include food substances like wheat, egg, milk, and chocolate; inhalants like pollen grains, fungi, dust, and smoke; contactants like chemical substances, metals, animals, and plants; infectious agents like parasites, bacteria, viruses, and fungi; drugs like aspirin and antibiotics; and physical agents like cold, heat, light, pressure, and radiation.
It is important to note that the terms “allergy” and “hypersensitivity” are often used interchangeably. However, an allergic reaction typically refers to the signs and symptoms that an animal experiences. In contrast, a hypersensitivity reaction describes the immune response itself.
Gabriel, S., Geldhof, P., Phiri, I. K., Cornillie, P., Goddeeris, B. M., & Vercruysse, J. (2005). Placental transfer of immunoglobulins in cattle infected with Schistosoma mattheei. Veterinary Immunology and Immunopathology, 104(3-4), 265-272.
Gattinoni, L., Speiser, D. E., Lichterfeld, M., & Bonini, C. (2017). T memory stem cells in health and disease. Nature Medicine, 23(1), 18-27.
Hall, J. E., & Hall, M. E. (2020). Guyton and hall textbook of medical physiology e-Book. Elsevier Health Sciences.
Klein, T. B. G. (2012). Cunningham’s textbook of veterinary physiology-E-book. Elsevier Health Sciences.
Lissoni, P., Messina, G., Pelizzoni, F., Rovelli, F., Brivio, F., Monzon, A.,... Di Fede, G. (2020). The fascination of cytokine immunological science. Journal of Infectiology and Epidemiology, 3(1), 18-28.
Louveau, A., Plog, B. A., Antila, S., Alitalo, K., Nedergaard, M., & Kipnis, J. (2017). Understanding the functions and relationships of the glymphatic system and meningeal lymphatics. The Journal of Clinical Investigation, 127(9), 3210-3219.
Tizard, I. R. (2017). Veterinary immunology-e-book. Elsevier Health Sciences.
von der Weid, P. Y., & Muthuchamy, M. (2010). Regulatory mechanisms in lymphatic vessel contraction under normal and inflammatory conditions. Pathophysiology, 17(4), 263-276.
Wiig, H., & Swartz, M. A. (2012). Interstitial fluid and lymph formation and transport: Physiological regulation and roles in inflammation and cancer. Physiological Reviews, 92(3), 1005-1060.
More on the topic IMMUNITY:
- EPIDEMIOLOGY
- Control
- Smith Bradford P., Van Metre David C., Pusterla Nicola (eds.). Large Animal Internal Medicine. Part 2. 6th edition. — Elsevier,2020. — 2279 p., 2020
- Smith Mary C., Sherman David M.. Goat Medicine. 3rd edition. — Wiley-Blackwell,2023. — 976 p., 2023
- Vogelnest L., Portas T. (Eds.). Current Therapy in Medicine of Australian Mammals. CSIRO,2025. — 848 p., 2025
- Interdigital Necrobacillosis (Foot Rot) in Cattle
- Diseases Caused by Bovine Viral Diarrhea Virus (BVDV)
- THE PAEDIATRIC CONSULTATION AND CLINICAL CONSIDERATIONS
- Mycoplasma Polyarthritis in Goats
- References